
Вирусы как инфекционные агенты являются
Вирусы являются внутриклеточными инфекционными агентами. Весь репликативный цикл вируса осуществляется с исполь зованием метаболических и генетических ресурсов клеток. Поэтому патогенез вирусных инфекций, в первую очередь, следует рассматривать на молекулярном и клеточном уровнях (В.И. Покровский, О.И. Киселев, 2002). Вместе с тем, инфекционный процесс, вызванный вирусами, развивается в пределах того или иного органа или ткани, так как большинство вирусов обладают достаточно высокой органной или тканевой тропностью. Поэтому характер развития внутритканевых процес сов при вирусных инфекциях, с одной стороны, определяется, как правило, цитопатическим действием вируса на клетки данной ткани и органа, а с другой стороны, реакцией внутритканевых и органных систем защиты от вирусной инфекции. Последние не редко носят деструктивный характер и усугубляют течение забо левания и его последствия. Поэтому понимание патогенеза вирусных инфекций представляется исключительно важным для создания рациональных схем лечения, основанных на гармоничном сочетании симптоматического лечения и противовирусной химиотерапии.
Стадии репликативного цикла вируса гриппа представлены на рис. 1. Такой же репликативный цикл свойствен большинству РНК- и ДНК-содержащих вирусов, кроме тех, кото рые обладают особыми механизмами хронизации процесса, латенции и способностью к интеграции в клеточный геном (например, ВИЧ, гепатит С, вирус папилломы и.т.). Ключевыми стадиями репликативного цикла вируса являются проникновение в клетки, декапсидация, активация процессов транскрипции и трансляции, собственно репликация вирусного генома и созревание зрелых вирусных частиц с выходом нового потомства инфекционного вируса.
Проникновение вируса в клетки - одна из ключевых стадий патогенеза инфекционного процесса. Для большинства вирусов степень цитопатического действия вируса прямо зависит от множественности заражения. В естественных условиях инфицирова ние клеток вирусами не происходит обычно с высокой множе ственностью. Исключение составляют респираторные вирусы, которые передаются воздушно-капельным путем с высокой инфицирующей дозой. ВИЧ и гепатиты, наоборот, эффективно передаются с кровью в самых незначительных трудно определяе мых титрах. Такие вирусы имеют особые свойства, определяющие их контагиозность.
Для проникновения вируса в клетку, необходимо: высокое сродство к вирусоспецифическому рецептору; множественность рецепторов; эффективность слияния вируса с клетками при инфицировании; способность к образованию синцития, к образованию гигантских синпластов, состоящих из множества слившихся клеток, в которых вирусный нуклеопротеид без эндоцитоза беспрепятственно переходит от одной зараженной клетки к множеству других через цитоплазму.
Такими свойствами из респираторных вирусов обладает респираторно-синцитиальный вирус, а среди ретровирусов – вирус иммунодефицита человека.
Стадии жизненного цикла вируса гриппа и соответствующие ингибиторы репродукции в известной степени могут быть экстраполированы на многие другие вирусы, за исключением тех, жизненный цикл которых включает интеграцию в клеточный геном. В процессе репликации вирусов обычно происходит селективная блокада трансляции клеточных мРНК и активация экспресси клеточных генов, кодирующих провоспапительные лимфокины. Многократные раунды репликации вирусного генома приводят к истощению энер гетических ресурсов клеток и пула предшественников азотистых оснований. Повышенное содержание провоспалительных лимфокинов приводит к развитию воспалительной реакции и локально му генерированию свободных радикалов, оказывающих, в свою очередь, повреждающее действие на клетки и ткани через перекисное окисление липидов клеточных мембран, инактивацию жизненно важных ферментов. Поэтому одним из важнейших факторов патогенности вирусов является их репликативный потенциал: чем активнее происходит репликация, тем сильнее цитопатическое действие вирусов на клетки и пораженную ткань (орган).
Значительный вклад в цитопатическое действие вирусов вносит репродукция вирусов в митохондриях, что сопровождается нарушением функций этих органоидов, падением продукции АТФ и индукцией опосредованного митохондриями пути апоптоза.
Впервые возможность репродукции вирусов в митохондриях была доказана выдающимся отечественным вирусологом В.М. Ждановым. Узкий тропизм вирусов, в известной степени, спасителен для организма, так как инфекционный процесс ограничен одним органом.
Это обстоятельство позволяет, проводит рациональную симптоматическую терапию, направленную на защиту данной ткани или органа и ориентировать противовирусные средства в данный орган, например, легкие.
Возбудители гриппа относятся к семейству ортомиксовирусов (Orthomyxo-viridae) и являются пневмотропными вирусами. Схематическое строение вируса гриппа представлено на рис. 2.1. Геном вируса складывается из 8-ми фрагментов однонитчатой РНК, которые кодируют
10 вирусных белков. Фрагменты РНК имеют общую белковую оболочку, соединяющую их, образуя антигенно-стабильный рибонуклеопротеид (S-антиген), который определяет принадлежность вируса к серотипу А, В или С. Снаружи вирус покрыт двойным липидным слоем с внутренней стороны которого находится слой мембранного белка.
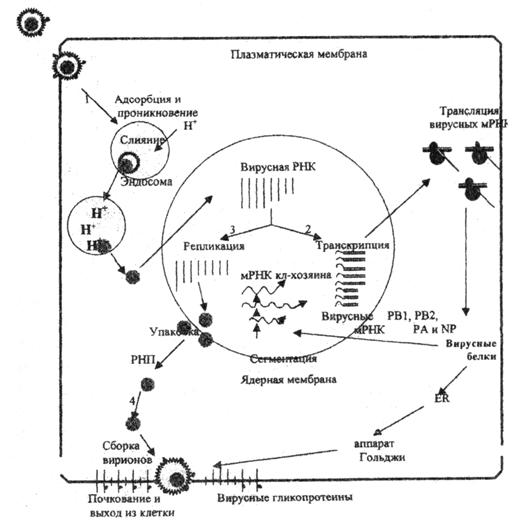
Рис. 1. Репликативный цикл вируса гриппа (В.И.Покровский, О.И.Киселев, 2002)
Представлены 4 основные фазы репликативного цикла вируса гриппа: 1 – адсорбция и проникновение вируса в клетку хозяина; 2 – транскрипция вирусной РНК и трансляция вирусных белков; 3 – репликация вирусной РНК и 4 – самосборка вирионов и последующий выход их из клетки.
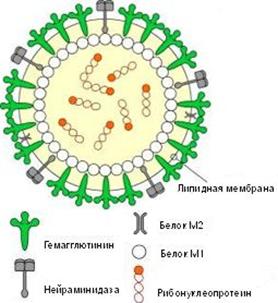
Рис. 2. Схематическое строение вируса гриппа
Гемаглютинин и нейраминидаза являются факторами агрессии вируса гриппа. Интенсивность интоксикации при гриппе определяется свойствами гемаглютинина, а нейраминидаза обладает выраженным иммунодепресивным действием. Оба поверхностных антигена характеризуются выраженной способностью к изменчивости, в результате чего появляются новые антигенные варианты вируса. Гемаглютинины 1, 2, 3 типов и нейраминидазы 1, 2 типов содержат вирусы, которые поражают человека. Другие антигены характерные для вирусов гриппа животных (свиней, собак, лошадей, многих видов птиц и др.).
Вирусы гриппа А. Эти возбудители более вирулентны и контагиозны, чем вирусы гриппа В и С. Это обусловлено тем, что вирус А содержит 2 типа нейраминидазы (N1, N2) и 4 –гемаглютинина (H0, H1, H2, H3); вирус В – 1 и 1 соответственно, поэтому он имеет лишь антигенные варианты внутри одного серотипа; вирус С содержит только гемаглютинин и не содержитнейраминидазу, но он имеет рецептор, который разрушает энзим. Вирусу С не свойственная изменчивость.
Одновременно циркулируют и имеют эпидемическое распространение 2 подтипа вируса гриппа А H3N2 и H1N1. На сегодняшний день во всем мире доминирует вариант А/Сидней/05/97, А/Берн/07/95 и А/Пекин/262/95. Характерной чертой современных вирусов гриппа А (H3N2) является изменение ряда их биологических свойств: тяжело размножаются в куриных эмбрионах, не агглютинируют эритроциты кур, взаимодействуют только с эритроцитами человека или морской свинки, имеют низкую иммуногенную активность.
Вирусы гриппа В выделяются только от людей. По вирулентности и эпидемиологической значимости они уступают вирусам гриппа А. По антигенным свойствам гемаглютинин инейраминидазу вируса В можно разделить на 5 подтипов. Их изменчивость носит более постепенный и медленный характер, что объясняет эпидемиологические особенности вируса гриппа В (эпидемии возникали 1 раз в 3-4 года до 1988 г.). В мире сейчас циркулируют 2 варианта вируса гриппа В: В/Пекин/184/93 (Европа, Америка, Африка, Австралия) и В/Виктория/2/87 (Юго-Восточная Азия). Характерной чертой современных вирусов гриппа А и В является их высокая чувствительность к ингибиторам гемаглютинирующей и инфекционной активности, которые содержатся в сыворотке крови нормальных животных.
Вирусы гриппа С . Вирусы гриппа С, в отличие от вирусов А и В, не вызывают эпидемий, но они могут быть причиной вспышек в организованных детских коллективах. Подъемы заболеваемости гриппом С нередко предшествуют или сопровождают эпидемии гриппа А и В. У детей раннего возраста и более старших возрастных группах он приводит к заболеваниям в легкой ибессимптомной форме.
Для вирусов гриппа С характерна значительно большая стабильность антигенных и биологических свойств. По ряду биологических характеристик эти возбудители отличаются от других представителей семейства ортомиксовирусов. Им присуща низкая репродуктивная активность в разных клеточных системах и наличие других, чем у вирусов гриппа А и В, рецепторов на поверхности эритроцитов. Рецептор-деструктирующая активность связана не с нейраминидазой, как у вирусов гриппа А и В, а с ферментом нейраминат-0-ацетилестеразою.
Сложность этиологии усиливают факты выделения от людей вирусов, которые не свойственные человеку. Гены вируса гриппа (H5N1), выделенные в 1997 году от человека в Гонконге, оказались подобными генам птичьего вируса, который преодолел межвидовой барьер. Оказалось, что человеческие и птичьи вирусы гриппа были реасортантами (рекомбинантные штаммы), которые получили внутренние гены РВ1 и РВ2 от перепелиного вируса гриппа А (H9N2) – штаммы А/перепел/Гонконг/61/97.
Вирусы гриппа малоустойчивы во внешней среде, в течение нескольких часов при комнатной температуре разрушаются. Под действием дезинфицирующих растворов (спирта, формалина, сулемы, кислот, щелочей) они быстро погибают. Нагревание до 50-60°С инактивирует вирусы в течение нескольких минут, в замороженном состоянии при температуре -70° С они сохраняются годами, не теряя инфекционных свойств, быстро погибают под воздействием ультрафиолетового излучения. Оптимум размножения вируса происходит при +37°С в слабощелочной среде. Как и другие вирусы, возбудитель гриппа отнюдь не чувствительный к антибиотикам и сульфаниламидам.
Вирус гриппа не растет на обычных питательных средах. Его можно выделить из материала, который получен от больного в первые дни болезни (смывы из носоглотки, мокроты), путем заражения культур или клеток куриных эмбрионов.
Традиционно считалось, что человеческий организм освобождается от возбудителя в ближайшие дни после выздоровления. Однако постепенно накапливались косвенные свидетельства, которые опровергают эту точку зрения. И в 1985 г. были получены прямые доказательства формирования длительной персистенции вируса гриппа. У одного человека удалось многократно выделять вирус гриппа в течение 9-ти месяцев и дольше. Возможность пожизненной персистенции вируса в организме человека продолжает изучаться.
1. Грипп и другие респираторные вирусные инфекции / под ред. О.И. Киселева, И.Г. Мариничева, А.А. Сомининой. – СПб. – 2003.
2. Дриневский В.П., Осидак Л.В., Цыбалова Л.М. Острые респираторные инфекции у детей и подростков // Практическое руководство под редакцией О.И. Киселева. – Санкт-Петербург. – 2003.
3. Железникова Г.Ф., Иванова В.В., Монахова Н.Е. Варианты иммунопатогенеза острых инфекций у детей. – Санкт-Петербург. – 2007. – 254 с.
4. Ершов Ф.И. Грипп и другие ОРВИ // Антивирусные препараты. Справочник. – М., 2006. –С. 226-247.
5. Ершов Ф.И., Романцов М.Г. Антивирусные средства в педиатрии. – М., 2005. – С.159-175.
6. Ершов Ф.И., Киселев О.И. Интерфероны и их индукторы (от молекул до лекарств). – М., 2005. – С. 287-292.
7. Иванова В.В. Острые респираторно-вирусные заболевания // Инфекционные болезни у детей. – М., 2002.
8. Онищенко Г.Г., Киселев О.И., Соминина А.А. Усиление надзора и контроля за гриппом как важнейший элемент подготовки к сезонным эпидемиям и очередной пандемии. – М., – 2004. – С.5-9.
9. Об утверждении стандарта медицинской помощи больным гриппом, вызванным идентифицированным вирусом гриппа (грипп птиц) // Приказ Минздравсоцразвития №460 от 07.06.2006 г.
10. Романцов М.Г., Ершов Ф.И. Часто болеющие дети: Современная фармакотерапия. – М., 2006. – 192 с.
11. Стандартизированные принципы диагностики, лечения и экстренной профилактики гриппа и других острых респираторных инфекций у детей / под ред. О.И. Киселева. – СПб. – 2004. – С. 82-95.
12. Лекарственные средства в фармакотерапии патологии клетки / под редакцией Т.Г. Кожока.-М., 2007.

В 1901 американский военный хирург У.Рид и его коллеги установили, что возбудитель желтой лихорадки также является фильтрующимся вирусом. Желтая лихорадка была первым заболеванием человека, опознанным как вирусное, однако потребовалось еще 26 лет, чтобы ее вирусное происхождение было окончательно доказано.
Свойства и происхождение вирусов.
Наиболее просто устроенные вирусы состоят из нуклеиновой кислоты, являющейся генетическим материалом (геномом) вируса, и покрывающего нуклеиновую кислоту белкового чехла. В состав некоторых вирусов входят также углеводы и жиры (липиды). Таким образом, вирусы можно рассматривать просто как мобильные наборы генетической информации. Вирусы лишены некоторых ферментов, необходимых для репродукции, и могут размножаться только внутри живой клетки, метаболизм которой после заражения перестраивается на воспроизводство вирусных, а не клеточных компонентов. Это свойство вирусов позволяет отнести их к облигатным (обязательным) клеточным паразитам. После синтеза отдельных компонентов формируются новые вирусные частицы. Симптомы вирусного заболевания развиваются как следствие повреждения вирусами отдельных клеток.
Принято считать, что вирусы произошли в результате обособления (автономизации) отдельных генетических элементов клетки, получивших, кроме того, способность передаваться от организма к организму. В нормальной клетке происходят перемещения нескольких типов генетических структур, например матричной, или информационной, РНК (мРНК), транспозонов, интронов, плазмид. Такие мобильные элементы, возможно, были предшественниками, или прародителями, вирусов.
Являются ли вирусы живыми организмами?
В 1935 американский биохимик У.Стэнли выделил в кристаллической форме вирус табачной мозаики, доказав тем самым его молекулярную природу. Полученные результаты вызвали бурные дискуссии о природе вирусов: являются ли они живыми организмами или просто активированными молекулами? Действительно, внутри зараженной клетки вирусы проявляют себя как интегральные компоненты более сложных живых систем, но вне клетки представляют собой метаболически инертные нуклеопротеины. Вирусы содержат генетическую информацию, но не могут самостоятельно реализовать ее, не обладая собственным механизмом синтеза белка. Когда особенности строения и репродукции вирусов оказались выясненными, вопрос о том, являются ли они живыми, постепенно утратил свое значение.
Размеры вирусов.
Величина вирусов варьирует от 20 до 300 нм (1 нм = 10 -9 м). Практически все вирусы по своим размерам мельче, чем бактерии (см. БАКТЕРИИ). Однако наиболее крупные вирусы, например вирус коровьей оспы, имеют такие же размеры, как и наиболее мелкие бактерии (хламидии и риккетсии), которые тоже являются облигатными паразитами и размножаются только в живых клетках. Поэтому отличительными чертами вирусов по сравнению с другими микроскопическими возбудителями инфекций служат не размеры или обязательный паразитизм, а особенности строения и уникальные механизмы репликации (воспроизведения самих себя).
СТРОЕНИЕ ВИРУСОВ
Вирионы со спиральным типом симметрии, как у вируса табачной мозаики, имеют форму удлиненного цилиндра; внутри белкового чехла, состоящего из отдельных субъединиц – капсомеров, находится свернутая спираль нуклеиновой кислоты (РНК). Вирионы с икосаэдрическим типом симметрии (от греч. eikosi – двадцать, hedra – поверхность), как у полиовируса, имеют сферическую, а точнее, многогранную форму; их капсиды построены из 20 правильных треугольных фасеток (поверхностей) и похожи на геодезический купол.
Встречаются вирусы с еще более сложным строением. Вирионы поксвирусов (вирусы группы оспы) не имеют правильного, типичного капсида: между сердцевиной и наружной оболочкой у них располагаются трубчатые и мембранные структуры.
РЕПЛИКАЦИЯ ВИРУСОВ
ДНК обычно существует в виде двухцепочечных структур: две полинуклеотидные цепочки соединены водородными связями и закручены таким образом, что образуется двойная спираль. РНК, напротив, обычно существует в виде одноцепочечных структур. Однако геном отдельных вирусов представляет собой одноцепочечную ДНК или двухцепочечную РНК. Нити (цепочки) вирусной нуклеиновой кислоты, двойные или одинарные, могут иметь линейную форму или замыкаться в кольцо.
У некоторых ДНК-содержащих вирусов сам цикл репродукции в клетке не связан с немедленной репликацией вирусной ДНК; вместо этого вирусная ДНК встраивается (интегрируется) в ДНК клетки-хозяина. На этой стадии вирус как единое структурное образование исчезает: его геном становится частью генетического аппарата клетки и даже реплицируется в составе клеточной ДНК во время деления клетки. Однако впоследствии, иногда через много лет, вирус может появиться вновь – запускается механизм синтеза вирусных белков, которые, объединяясь с вирусной ДНК, формируют новые вирионы.
Так называемые ретровирусы содержат в качестве генома РНК и имеют необычный способ транскрипции генетического материала: вместо транскрипции ДНК в РНК, как это происходит в клетке и характерно для ДНК-содержащих вирусов, их РНК транскрибируется в ДНК. Двухцепочечная ДНК вируса затем встраивается в хромосомную ДНК клетки. На матрице такой вирусной ДНК синтезируется новая вирусная РНК, которая, как и другие, определяет синтез вирусных белков. См. также РЕТРОВИРУСЫ.
КЛАССИФИКАЦИЯ ВИРУСОВ
Тем не менее система классификации вирусов необходима в практической работе, и попытки ее создания предпринимались неоднократно. Наиболее продуктивным оказался подход, основанный на структурно-функциональной характеристике вирусов: чтобы отличить разные группы вирусов друг от друга, описывают тип их нуклеиновой кислоты (ДНК или РНК, каждая из которых может быть одноцепочечной или двухцепочечной), ее размеры (число нуклеотидов в цепочке нуклеиновой кислоты), число молекул нуклеиновой кислоты в одном вирионе, геометрию вириона и особенности строения капсида и наружной оболочки вириона, тип хозяина (растения, бактерии, насекомые, млекопитающие и т.д.), особенности вызываемой вирусами патологии (симптомы и характер заболевания), антигенные свойства вирусных белков и особенности реакции иммунной системы организма на внедрение вируса.
В систему классификации вирусов не вполне укладывается группа микроскопических возбудителей болезней, называемая вироидами (т.е. вирусоподобными частицами). Вироиды вызывают многие распространенные среди растений болезни. Это мельчайшие инфекционные агенты, лишенные даже простейшего белкового чехла (имеющегося у всех вирусов); они состоят только из замкнутой в кольцо одноцепочечной РНК.
ВИРУСНЫЕ ЗАБОЛЕВАНИЯ
Хотя вирусы не являются полноценными живыми организмами, их эволюционное развитие имеет много общего с эволюцией других патогенных организмов. Для того чтобы сохраниться как вид, ни один паразит не может быть слишком опасным для своего основного хозяина, в котором размножается. В противном случае это привело бы к полному исчезновению хозяина как биологического вида, а вместе с ним и самого возбудителя. В то же время любой патогенный организм не сможет существовать как биологический вид, если у его основного хозяина слишком быстро и эффективно развивается иммунитет, позволяющий подавлять репродукцию возбудителя. Поэтому вирус, вызывающий острое и тяжелое заболевание у какого-либо вида животных, обычно имеет еще и другого хозяина. Размножаясь в последнем, вирус не наносит ему (как виду) существенного вреда, однако такое относительно безвредное сосуществование поддерживает циркуляцию вируса в природе. Так, например, вирус бешенства в природе сохраняется среди грызунов, для которых заражение этим вирусом не является смертельным.
Природным резервуаром для вирусов лошадиных энцефалитов, особо опасных для лошадей и в несколько меньшей степени для человека, являются птицы. Эти вирусы переносятся кровососущими комарами, в которых вирус размножается без существенного вреда для комара. Иногда вирусы могут передаваться насекомыми пассивно (без размножения в них), однако чаще всего они репродуцируются в переносчиках.
Для многих вирусов, например кори, герпеса и отчасти гриппа, основным природным резервуаром является человек. Передача этих вирусов происходит воздушно-капельным или контактным путем.
Распространение некоторых вирусных заболеваний, как и других инфекций, полно неожиданностей. Например, в группах людей, проживающих в антисанитарных условиях, практически все дети в раннем возрасте переносят полиомиелит, обычно протекающий в легкой форме, и приобретают иммунитет. Если же условия жизни в этих группах улучшаются, дети младшего возраста обычно полиомиелитом не болеют, но заболевание может возникнуть в более старшем возрасте, и тогда оно часто протекает в тяжелой форме.
Возбудители некоторых болезней, в том числе очень тяжелых, не укладываются ни в одну из вышеперечисленных категорий. К особой группе медленных вирусных инфекций еще недавно относили, например, болезнь Крейтцфельда – Якоба и куру – дегенеративные заболевания головного мозга, имеющие очень продолжительный инкубационный период. Однако оказалось, что они вызываются не вирусами, а мельчайшими инфекционными агентами белковой природы – прионами (см. ПРИОН).
Репродукция вирусов тесно переплетается с механизмами синтеза белка и нуклеиновых кислот клетки в зараженном организме. Поэтому создать лекарства, избирательно подавляющие вирус, но не наносящие вреда организму, – задача чрезвычайно трудная. Все же оказалось, что у наиболее крупных вирусов герпеса и оспы геномные ДНК кодируют большое число ферментов, отличающихся по свойствам от сходных клеточных ферментов, и это послужило основой для разработки противовирусных препаратов. Действительно, создано несколько препаратов, механизм действия которых основан на подавлении синтеза вирусных ДНК. Некоторые соединения, слишком токсичные для общего применения (внутривенно или через рот), годятся для местного использования, например при поражении глаз вирусом герпеса.
Известно, что в организме человека вырабатываются особые белки – интерфероны. Они подавляют трансляцию вирусных нуклеиновых кислот и таким образом угнетают размножение вируса. Благодаря генной инженерии стали доступны и проходят проверку в медицинской практике интерфероны, производимые бактериями (см. ГЕННАЯ ИНЖЕНЕРИЯ).
К самым действенным элементам естественной защиты организма относятся специфические антитела (специальные белки, вырабатываемые иммунной системой), которые взаимодействуют с соответствующим вирусом и тем самым эффективно препятствуют развитию болезни; однако они не могут нейтрализовать вирус, уже проникший в клетку. Примером может служить герпетическая инфекция: вирус герпеса сохраняется в клетках нервных узлов (ганглиев), где антитела не могут его достичь. Время от времени вирус активируется и вызывает рецидивы заболевания.
Обычно специфические антитела образуются в организме в результате проникновения в него возбудителя инфекции. Организму можно помочь, усиливая выработку антител искусственно, в том числе создавая иммунитет заранее, с помощью вакцинации. Именно таким способом, путем массовой вакцинации, заболевание натуральной оспой было практически ликвидировано во всем мире. См. также ВАКЦИНАЦИЯ И ИММУНИЗАЦИЯ.
Для приготовления вакцинных препаратов необходимо накопить вирус. С этой целью часто используют развивающиеся куриные эмбрионы, которых заражают данным вирусом. После инкубирования зараженных эмбрионов в течение определенного времени накопившийся в них вследствие размножения вирус собирают, очищают (центрифугированием или другим способом) и, если нужно, инактивируют. Очень важно удалить из препаратов вируса все балластные примеси, которые могут вызывать серьезные осложнения при вакцинации. Конечно, не менее важно убедиться, что в препаратах не осталось неинактивированного патогенного вируса. В последние годы для накопления вирусов широко используют различные типы клеточных культур.
МЕТОДЫ ИЗУЧЕНИЯ ВИРУСОВ
Работы с бактериофагами способствовали расширению методического арсенала в изучении вирусов животных. До этого исследования вирусов позвоночных выполнялись в основном на лабораторных животных; такие опыты были очень трудоемки, дороги и не очень информативны. Впоследствии появились новые методы, основанные на применении тканевых культур; бактериальные клетки, использовавшиеся в экспериментах с фагами, были заменены на клетки позвоночных. Однако для изучения механизмов развития вирусных заболеваний эксперименты на лабораторных животных очень важны и продолжают проводиться в настоящее время.
Не уверен, что меньше. Но проще – точно есть.
Помимо прионов, которые мельче вирусов и состоят только из белка, не имея нууклеиновой кислоты, есть ещё вироиды и вирусоиды – инфекционные агенты, состоящие только из одной "голой" кольцевой РНК, без какой-либо оболочки и белков. "Геном" такого агента меньше генома любого из вирусов. Но за линейные размеры поручиться не могу – линейный размер ДНК зависит от плотности её упаковки, а вироиды – весьма малоизученный класс патогенов. Скорее всего, линейкой их вряд ли кто-то мерил.
В основном они вызывают болезни растений.
Разница между вироидами и вирусоидами – в потребности в вирусе-"помощнике". Вироиды справляются со всем сами: им не нужна "одёжка"-капсид, а для репликации они используют ферменты клетки-хозяина и собственную каталитическую активность (напомню что некоторые РНК могут служить катализаторами, подобно ферментам – такие РНК называют рибозимами).
А вирусоиды – это РНК-патогены, которым для репликации необходимо присутствие в клетке другого вируса, полноценного. А "путешествует" вирусоид кнутри капсида полноценного вируса, вместе с его нуклеиновой кислотой. Поэтому вирусоиды называют ещё сателлитными РНК.
Любопытно, что у человека тоже известен такой патоген. Пока он один. Это вирус гепатита D, или дельта-агент. Он может оказать своё действие, только если человек уже заражён вирусом гепатита B. Наличие вируса гепатита D значительно утяжеляет течение заболевания, такие пациенты имеют больше шансы перейти в цирроз печени и погибнуть. Этот патоген для человека необычен, зато для растений – вполне обычная вещь. Есть теория, что данный патоген "перекинулся" на человека от растений, а ранее был обычно растительной сателлитной РНК.
Вирус гепатита D передаётся теми же путями, что его "помощник": половым путём и через кровь. Поэтому меры профилактики те же самые, что и для вируса гепатита B: использование одногоразового иструментария для инъекций; стерилизация медицинского, стоматологического, парикмахерского инструментария и инструментов для татуажа; отказ от употребления наркотиков; защищённый половой контакт.
Кроме того, прививка от гепатита B защищает и от гепатита D – если полноценный вирус не сможет размножиться, сателлитная РНК тоже безобидна. Поэтому в настоящее время рекомендуется прививаться от гепатита B, чтобы обезопасить себя.
А теперь внимание! Вспомните вирус иммунодефицита человека, который умеет "переписываться" с РНК на ДНК и обратно, в виде ДНК подолгу находясь в геноме клетки-хозяина? Он не единственный в своём роде, такие вирусы называются ретровирусами (от латинского "ретро-" – обратно). Представьте, себе "вироид", соответствующий ретровирусу. Это мобильные генетические элементы – транспозоны и ретротраснпозоны, и их много в нашем геноме.
Про них собрана целая подборка статей на нашем сайте "Биомолекула".
До половины всей нашей ДНК состоит из таких элементов. Они не способны сформировать инфекционнцю частицу и выйти из клетки, но копировать себя вунтри генома и перемещаться на новые места вполне способны (ваот и "расползлись" на половину генома). Такой "инфекционный агент для домашшего пользования". Многие из них произошли от ретровирусов, когда-то заразивших наших предков и впоследствии утративших "оболочки" и активность.
Но они сыграли важную роль в эволюции: подарили нам способность к обучению и памяти (статья на Биомолекуле) и способность к формированию плаценты и вынашиванию потомства (ещё одна статья на Биомолекуле). Это вполне себе инфекционные элементы – но они тоже представляют из себя "голую" ДНК. Они меньше, чем любой вирус. Если бы они могли говорить, то вполне могли бы сказать о себе словами Волан-де-Морта:
"Я был вырван из тела, я стал меньше, чем дух, чем самое захудалое привидение. но всё-таки я был жив. Я, который дальше еого-либо прошёл по тропе, ведущей к бессмертию"
Так что инфекционные элементы, которые меньше вируса, существуют. И мы наполовину состоим из них.
А вот есть ли что-то инфекционное меньше вироида и приона. Скорее всего, на Земле нет. Чтобы "записать" наследственную информацию, нужна какая-нибудь биомакромолекула – нуклеиновая кислота или белок. В химии и физике известны цепные реакции, которые чем-то напоминают процесс распространения инфекции. Но такие системы не считаются живыми, поэтому, вероятно, ниже вироидов, вирусоидов и прионов мир инфекций заканичвается. Хотя.. кто знает?
, PhD, Wadsworth Center, NYSDOH
Last full review/revision February 2018 by Laura D Kramer, PhD
Вирусы – это мельчайшие паразиты, обычно величиной от 0,02 до 0,3 μ м, хотя недавно были обнаружены несколько очень крупных вирусов длиной до 1 μ м (megavirus, pandoravirus). Размножение вирусов находится в полной зависимости от клеток (бактериальных, растительных или животных). Вирусы имеют внешнюю белковую, а иногда и липидную, оболочку, ядро РНК или ДНК и иногда ферменты, необходимые для первых этапов репликации вируса.
Вирусы классифицируются преимущественно в соответствии с природой и структурой их генома и способом их репликации, а не в зависимости от заболеваний, которые они вызывают. Таким образом, существуют ДНК-вирусы и РНК-вирусы; каждый тип может иметь одинарные или двойные цепи генетического материала. Одноцепочечные РНК-вирусы в свою очередь подразделяются на РНК с положительной полярностью и РНК с отрицательной полярностью. Как правило, ДНК-вирусы реплицируются в ядре клетки-хозяина, а РНК-вирусы обычно реплицируются в цитоплазме. В то же время, некоторые одноцепочечные РНК-вирусы с положительной полярностью, называемые ретровирусами, используют совершенно другой способ репликации.
Ретровирусы используют обратную транскрипцию для создания двухцепочечной ДНК-копии (провируса) их генома РНК, которая встраивается в геном клетки-хозяина. Обратная транскрипция осуществляется с помощью фермента обратной транскриптазы, который вирус несет с собой внутри своей оболочки. Примерами ретровирусов являются вирусы иммунодефицита человека и вирусы Т-клеточного лейкоза человека. После того, как провирус интегрируется в ДНК клетки-хозяина, он транскрибируется с использованием обычных клеточных механизмов для воспроизведения вирусных белков и генетического материала. Если инфицированная клетка относится к зародышевой линии, интегрированный провирус может укорениться как эндогенный ретровирус, который передаётся потомству.
Секвенирование генома человека выявило, что как минимум, 1% генома человека состоит из эндогенных ретровирусных последовательностей, представляющие собой предыдущие контакты с ретровирусами в процессе эволюции человека. Немногие эндогенные ретровирусы человека остаются транскрипционно активными и вырабатывают функциональные белки (например, синтицины, которые участвуют в образовании структуры человеческой плаценты). Ряд экспертов полагает, что некоторые заболевания неясной этиологии, такие как рассеянный склероз, определенные аутоиммунные нарушения, а также различные злокачественные опухоли, могут быть вызваны эндогенными ретровирусами.
Поскольку транскрипция РНК не использует те же самые механизмы для контроля ошибок, что и транскрипция ДНК, РНК-вирусы – в частности, ретровирусы – особенно склонны к мутации.
Для того, чтобы произошло инфицирование, вирус в первую очередь прикрепляется к одиночной или к одной из нескольких рецепторных молекул на поверхности клетки-хозяина. Затем вирусная ДНК или РНК проникает в клетку-хозяин и отделяется от внешней оболочки (декапсуляция вируса) и воспроизводится в клетке-хозяине с участием определенных ферментов. Вновь синтезированные компоненты вируса затем собираются в полноценные вирусные частицы. Клетка–хозяин, как правило, погибает, выделяя новые вирусы, которые заражают другие клетки хозяина. Каждый этап вирусной репликации задействует различные ферменты и субстраты и дает возможность для противодействия процессу инфицирования.
Последствия вирусного инфицирования в значительной степени варьируются. Многие инфекции вызывают острое заболевание после непродолжительного инкубационного периода, а некоторые являются бессимптомными или вызывают несущественные симптомы, которые никак нельзя распознать, кроме как ретроспективно. Многие вирусные инфекции устраняются защитными силами организма, но некоторые переходят в латентную форму, а некоторые становятся причиной хронического заболевания.
При скрытой инфекции вирусная РНК или ДНК остается в клетках-хозяина, но не реплицируется и не вызывает заболеваний в течение долгого времени, иногда в течение многих лет. Латентные вирусные инфекции могут передаваться в течение бессимптомного периода, что облегчает распространение от человека к человеку. Иногда реактивацию вызывает инициирующий фактор (в частности, иммуносупрессия).
К распространённым вирусам, которые переходят в латентную форму, относятся
Читайте также:
