Основные структуры вируса фага т-4







480 руб. | 150 грн. | 7,5 долл. ', MOUSEOFF, FGCOLOR, '#FFFFCC',BGCOLOR, '#393939');" onMouseOut="return nd();"> Диссертация, - 480 руб., доставка 1-3 часа, с 10-19 (Московское время), кроме воскресенья
Автореферат - бесплатно , доставка 10 минут , круглосуточно, без выходных и праздников
Костюченко Виктор Анатольевич. Структура аппарата инфицирования бактериофага T4 : Дис. . канд. биол. наук : 03.00.03 : Москва, 2004 105 c. РГБ ОД, 61:04-3/1488
Введение к работе
Знание структуры вирусов и особенностей их формирования необходимо для разработки новых эффективных подходов в борьбе с вирусными инфекциями. Вирусы также являются удобной экспериментальной модельной системой для изучения механизмов ассоциации белковых субъединиц, которая может помочь в понимании процессов формирования сложных биологических структур клетки, таких как ядерные поры, центриоли, жгутики, компоненты мышечных филаментов и др
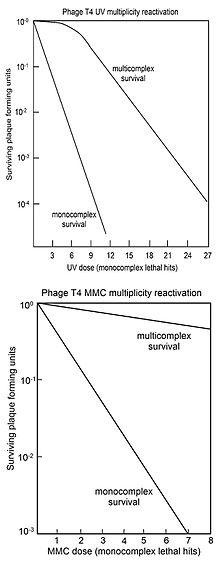
Бактериофаг Т4, инфицирующий Escherichia col:, является одним из основных модельных объектов молекулярной и структурной биологии [7] Частица фага Т4 (Рис I) состоит из двух основных частей — капсида ("головки"), содержащего геномную ДНК размером около 172 тысяч пар оснований, и "хвоста", который присоединен к капсиду через специальный портал на одной из вершин с симметрией 5-го порядка. Капсид является икосаэдром, удлинённым вдоль оси 5 порядка [5], имеет размеры 860 А в поперечнике и 1200 А длиной, и построен из 930 копий белка продукта гена (пг) 23, 55 копий пг24, 155 копий белка hoc ("highly antigenic outer capsid protein") и 810 копий белка soc ("small outer capsid protein") Портал содержит 12 копий белка пг20 Сократимый хвост состоит из трех частей "шейки", хвостового стержня, на котором собран сократимый хвостовой чехол, и базальной пластинки К шейке, построенной из гексамеров пг15, пг13 и/или пг14, присоединены "воротничок" и "бакенбарды", состоящие из тримеров белка фибритина (nrwac), воротничок, возможно, содержит также один из белков пг13 или пг14 Основную часть хвоста занимает сократимый хвостовой чехол и расположенный внутри него хвостовой стержень Чехол построен из 138 копий пг18, стержень состоит из пг!9, которого, видимо, 138 копий Завершает хвост базальная пластинка — многокомпо-
rife ^4^ Baseplate
Рис. 1: Бактериофаг Т4 .
а) криоэлектронная микрофотография; б) схематическая модель Рисунок взят из [19].
нентная молекулярная структура диаметром около 520 А и высотой в 270 А, состоящая из 16 различных белков, почти каждый из которых является олигомером. В состав базальной пластинки входятпг5, пгб, пг7, пг8, пг9, nrlO, nrl 1, пг12, пг25, пг26, пг27, пг28, пг29, пг48, пг53, и пг54. К базальной пластинке присоединены длинные фибриллы, построенные из пг34, пг35, пгЗб и пг37. Таким образом, в сборке частицы фага участвуют более тридцати различных белков, и это не не считая белков, связанных с ДНК внутри капсида, вспомогательных скаффолд-белков и белков-шаперонов.
С помощью электронной микроскопии с применением негативного контрастирования ранее удалось представить общее строение вируса. Хвостовой чехол бактериофага Т4 в растянутом (нативном) состоянии явился первой биологической структурой, для которой была проведена трёхмерная реконструкция [20]. Также была выполнена трёхмерная реконструкция сокращённого [1] хвоста и предложена двумерная модель базальной пластинки [4]. Разрешение этих моделей было довольно низким, около 40 А. Получить с высоким разрешением пространственную реконструкцию вирусной частицы целиком удавалось, в основном, лишь для сравнительно простых в устройстве икосаэдрических вирусов, поскольку отсутствовали необходимые вычислительные мощности и не были до конца разработаны методы реконструкции. В настоящее время, с совершенствованием методов электронной криомикроскопии и развитием компьютерных технологий, цель реконструировать частицу бактериофага Т4 с околоатомным разрешением представляется осуществимой.
Пространственная реконструкция такого сложного и большого вируса, каким является бактериофаг Т4 (размеры вируса примерно 2500 х 1000 х 1000 А, молекулярная масса частицы около 300 МДа) требует особого подхода, поскольку даже при использовании современной вычислительной техники необходимый объём данных и вычислений выходит за рамки осуществимого с помощью стандартных методов и программ. Поэтому, в данной работе были поставлены следующие задачи:
Разработать компьютерную программу для пространственной реконструкции больших объектов с различной симметрией по изображениям электронной криомикроскопии;
Провести пространственную реконструкцию комплекса базальной пластинки и хвостового стержня бактериофага Т4 (масса комплекса 8,8 МДа) и определить расположение и взаимодействие в нём белковых субъединиц, атомная структура которых ранее определена в нашей лаборатории: пг5, пг9, nrl 1, пг12 (фрагмент), пг27, атак же определить вероятную форму и расположение остальных белковых составляющих базальной пластинки;
На основании полученных результатов предложить механизм действия аппарата инфицирования бактериофага Т4, включающего хвостовой чехол и базалъную пластинку.
Структура и объём диссертации
Диссертация состоит из введения, четырёх глав, выводов, заключения и списка литературы. Объем диссертации составляет 105 страниц, включая 27 рисунков и 3 таблицы. Список цитируемой литературы содержит ПО наименований.
Научная новизна и практическая значимость
Впервые получена детальная пространственная картина важного функционального компонента сложного вируса — базальной пластинки бактериофага Т4, и предложено механистическое объяснение процесса заражения вирусом хозяйской клетки. Разработана компьютерная программа для получения реконструкций асимметричных, или имеющих низкую степень симметрии объектов, что позволит в будущем получить модели строения других вирусов и надмолекулярных биологических структур, представляющих научный и практический интерес. Полученные результаты и сведения структурного характера можно использовать, например, при конструировании молекулярных механизмов в нанотехнологии.
Результаты работы были доложены на Гордоновской конференции по дифракционным методам в биологии в США (Diffraction Methods in Molecular Biology Gordon Conference; Proctor Academy, Andovcr, New Hampshire, USA, 1998); на конференциях Медицинского института Говарда Хыоза в 2000, 2001, 2002 и 2004 гг (Howard Hughes Medial Institute Meeting of International Research Scholars, USA, 2000; Canada, 2001; Australia, 2002; Estonia, 2004); на конференции по сборке вирусов (XVIII Biennial Conference on Phage/Virus Assembly, USA, 2003).
По материалам диссертации опубликовано 12 печатных работ, из них 6 статей и 6 тезисов докладов.
Депонированные структурные данные
Депонированы в банк данных белковых структур PDB следующие структуры: пг5/пг27 - коды 1К28, 1PDJ, 1PDL; пг8 - коды 1N7Z, 1N80, 1N8B, 1PDM; пт9 - коды 1QEX, 1S2E, 1PDP; пгП - коды 1ЕГ6, IPDF; ПГІ2 - код 1PDI;
Реконструкция комплекса базальной пластинки и хвостового стержня депонирована в банк электронномикроскопических реконструкций (EMDB), код 1048.
Как протекает фаговая инфекция
Начинается все с того, что вирусная частица случайно сталкивается с клеткой бактерии. Фаг способен заразить вовсе не любую бактерию. Например, фаг, заражающий кишечную палочку, не замечает – сколько бы он с ними ни сталкивался – клетки стафилококка, и наоборот. Больше того, у той же кишечной палочки известно много разновидностей (их называют штаммами), и фаги, как правило, способны очень хорошо их различать. Они охотно заражают одни штаммы и совершенно игнорируют другие. Почему так происходит? Дело в том, что на поверхности бактериальной клетки есть структуры, к которым базальная пластинка фага по форме подходит, как ключ к замку. Такие структуры называются рецепторами. Есть рецептор для данного фага – фаг способен заражать эту бактерию, а эта бактерия способна стать его хозяином. Не подходит ключик? – Ну, стало быть, эта бактерия не для него.
Возникает вопрос, почему бактериальная клетка вынуждена жить под постоянной угрозой фаговой интервенции, не проще ли незаметно потерять где–то эти рецепторы и стать неприступной для фага? Но фаг тоже не прост, он использует в качестве причала структуры, предназначенные не для него, а совсем для других целей и жизненно важные для клетки, у которой, стало быть, есть веские причины не лишаться их.
Если фаг и бактерия узнали друг друга, фаг прочно связывается с ее поверхностью. Дальнейшая задача вируса состоит в том, чтобы ввести свой генетический материал внутрь бактериальной клетки, не нанеся ей – до поры до времени – вреда. Вот как это делает, например, фаг Т4, заражающий кишечную палочку – обычного обитателя нашего кишечника.
Бактериофаг Т4 является одним из наиболее сложно устроенных вирусов. Несколько вытянутая головка служит контейнером для хранения нуклеиновой кислоты. Сокращение чехла обеспечивает прободение бактериальной стенки. По внутреннему каналу стержневого отростка фаговая ДНК перемещается внутрь бактериальной клетки. К шестиугольной базальной пластинке крепятся длинные нити – фибриллы, которые обеспечивают начальный контакт фага с поверхностью бактерии. Чехол с одной стороны жестко закреплен на фаговой головке, а с другой прикреплен к базальной пластинке, находящейся на конце отростка. Чехол заметно короче отростка, поэтому, подобно пружине, пребывает в растянутом состоянии. После стыковки фага с бактерией форма базальной пластинки меняется, она больше не может удерживать чехол в растянутом состоянии, тот сжимается, и жесткий стержень как бы выстреливает из чехла, проминая клеточную стенку бактерии.

Бактериофаг Т4: 1 – головка; 2 – отросток, покрытый чехлом;3 – базальная пластинка; 4 – длинные фибриллы
Одного механического усилия недостаточно. Прогибаясь, клеточная стенка успешно сопротивляется попытке фагового отростка проткнуть ее, но оказывается бессильной противостоять секретному оружию фага – ферменту лизоциму, который находится вблизи острия отростка и, войдя в плотный контакт с клеточной стенкой, моментально прогрызает в ней дыру. Наконец–то стержень отростка пронзает клеточную стенку насквозь. Тут же по внутреннему каналу отростка, как сквозь игольное ушко, в бактерию впрыскивается нуклеиновая кислота, до того момента покоившаяся в фаговой головке.
На первый взгляд, задача может показаться не слишком сложной, но надо учесть, что молекула ДНК – очень длинная и чрезвычайно плотно упакована. Если уж сравнивать внутренний диаметр канала с игольным ушком, то в этом масштабе длина нити ДНК будет около метра, а скорость ее разматывания напоминает разматывание лески с катушки спиннинга при забрасывании блесны. И протащить нить надо так, чтобы не порвать ее. Порванная даже в одном месте, она уже неинфекционна и, стало быть, совершенно безвредна для бактерии и абсолютно бесполезна для фага.
Большинство фагов не имеют сократимого чехла, не говоря уже о том, что у многих и отростка–то никакого нет, ни большого, ни маленького. А преграда на пути серьезная – клеточная стенка бактерии. Она состоит из нескольких слоев и, подобно неповрежденной коже человека или кутикуле на поверхности листа, совершенно непроницаема для вируса. Как–то ее надо продырявить, и, так или иначе, все фаги умеют это делать.
Например, многие фаги связываются только с половыми пилями – нитевидными выростами на мужских клетках кишечной палочки. Да, как ни удивительно, у бактерий есть пол, мужской и женский, а у мужских клеток есть вырост, с помощью которого они конъюгируют с женской клеткой. Облепив вырост, фаги внедряют свою нуклеиновую кислоту в клетку бактерии. В общем, тем или иным способом, но фаги вводят свой генетический материал внутрь бактериальной клетки, оставляя на поверхности пустую, никому уже не нужную белковую оболочку.

Фаговая ДНК проникла внутрь бактерии. 1 – фаговая ДНК; 2 – стенка бактериальной клетки; 3 – сократившийся чехол; 4 – длинные фибриллы
С этого момента все меняется для бактериальной клетки. Вирус на время как бы исчезает. В клетке, куда проникла фаговая нуклеиновая кислота, не удается обнаружить никаких вирусных частиц. Более того, зараженная клетка выглядит совершенно нормальной. Но на самом деле жить ей осталось всего несколько минут. Под покровом клеточной стенки фаг начинает свое черное дело. Он заставляет все клеточные структуры работать на себя. Все ресурсы клетки отныне тратятся только на размножение фаговой ДНК, самой клетке уже ничего не достается. Белки образуются только фаговые, синтез клеточных компонентов совершенно подавлен или осуществляется лишь в той мере, в которой это нужно фагу. Многочисленные копии вновь образованной фаговой ДНК упаковываются в форме многогранника. Сверху они покрываются фаговым белком, и возникает зрелая фаговая головка с упакованной внутри нее ДНК. В другом месте клетки, в другом ее помещении налажено производство и сборка других фаговых белков, из которых образуется хвостовой отросток. Наконец, отростки и головки соединяются в полноценную фаговую частицу. Проникла в бактерию нуклеиновая кислота одной–единственной фаговой частицы, а теперь, через полчаса, их уже больше сотни. Им тесно, им пора покидать эту бактерию, с которой уже нечего взять. Вот только как это сделать? Бактерия мертва, но ее клеточная стенка все еще надежно удерживает взаперти многочисленное фаговое потомство.

Продольный разрез фага с пустой головкой: 1 – фибриллы, прикрепленные к головке;2 – "воротничок"; 3 – отросток;4 – канал, проходящий внутри отростка
И вновь приходит на помощь фаговый лизоцим. Он подгрызает клеточную стенку обреченной бактерии изнутри до тех пор, пока она достаточно не истончится и в конце концов не разорвется. Фаговое потомство выходит наружу и немедленно набрасывается на соседние бактерии, которые ожидает та же участь.
Такая инфекция называется продуктивной, а фаги, вызывающие продуктивную инфекцию – вирулентными.
| Вирус кишечной T4 | |
|---|---|
| классификация Вирус | |
| (Неоцениваемый): | Вирус |
| Тип: | incertae SEDIS |
| Учебный класс: | incertae SEDIS |
| Порядок: | Caudovirales |
| Семья: | Myoviridae |
| Род: | T4virus |
| Виды: | Вирус кишечной T4 |
| деформации | |
| |
| Синонимы | |