
Научные исследования бактерий и вирусов
Увеличивается во всем мире и число сторонников вирусной теории рака. Исследования сотен лабораторий свидетельствуют, что именно вирусы — наиболее вероятная причина рака, саркомы, лейкемии.
И. Губарев, наш специальный корреспондент, обратился к директору Института вирусологии имени И. Д. Ивановского АМН СССР, академику АМН СССР, профессору Виктору Михайловичу Жданову с просьбой рассказать об истории и сегодняшнем дне Вирусологии, о стратегии борьбы С вирусными болезнями.
Вирусология — наука молодая. 80 лет прошло со времени открытия И. Д. Ивановским первого вируса — возбудителя мозаичной болезни табака. Много позже — в 50-х годах — было получено первое несовершенное изображение этого инфекционного агента. Самые значительные исследования в области вирусологии были выполнены лишь за последние 15—20 лет.
С исследованиями вирусологов сегодня связано уничтожение инфекционных заболеваний на планете, борьба против рака. Вирусологии же, изучающей наиболее простые формы существования, предстоит дать ответ на многие вопросы, связанные с происхождением жизни на Земле.
Итак, что же мы знаем и «его еще не знаем о вирусах?
Пример: до недавнего времени мы почти ничего не знали о специфических обезьяньих вирусах. В 1960-х годах было начато массовое производство вакцины против полиомиелита, изготавливаемой на обезьяньих почках. Необходимо было обеспечить стерильность этой вакцины, то есть полностью исключить проникновение в нее каких-либо микроорганизмов. И вот в ходе исследований, направленных на обеспечение такого рода стерильности, был открыт целый ряд до тех пор неизвестных вирусов, специфичных для обезьян.
К настоящему времени мы располагаем сведениями примерно о тысяче видах вирусов. Безусловно, лучше других нам известны вирусы, поражающие человека. Их выявлено около 500 видов. Весьма обширна группа вирусов, найденных у лабораторных животных — мышей, кроликов, морских свинок.
Сравнительно много мы знаем о вирусах сельскохозяйственных животных и растений, меньше — о вирусах, опасных для птиц и других животных, древесных и кустарниковых пород лесе. И уж вовсе малоизвестны и числом и повадками вирусы папоротников, мхов, лишайников.
Вирусы проявляют себя не всегда одинаково. В одних случаях они нападают лишь на определенные виды живых существ. Скажем, уже выявлены специфические вирусы гриппа свиней, кошек, чаек, поражающие только этих животных и безопасные для других. Подчас специализация становится своеобразно утонченной: мельчайшие вирусы бактерий — фаги Р-17 выбирают в качестве объекта лишь мужские особи только одной разновидности кишечной палочки. А вот в числе объектов онкогенных вирусов — пресмыкающиеся, птицы, млекопитающие. Рекорд побивают, пожалуй, так называемые пулевидные вирусы, названные так благодаря их характерному очертанию на микрофотографии. Внешне вирусы этой разновидности очень схожи. А болезни они вызывают самые разнообразные, поражая при этом весьма далекие друг от друга виды живых существ. Они могут стать причиной бешенства — тяжелейшего поражения нервной системы млекопитающих (в том числе, разумеется, и человека) и таких болезней, как везикулярный стоматит крупного рогатого скота (передаваемый, кстати, через насекомых), желтой карликовости картофеля и полосатой штриховатости пшеницы. Эти же вирусы провоцируют тяжелое заболевание у мухи дрозофилы, приводящее насекомое к гибели в результате повышения чувствительности к углекислому газу.
Человек, животные, насекомые, растения. Болезни общие для многих видов и узко-специфичные. Откуда такой широкий спектр агрессивных возможностей? Под влиянием каких условий сложились эти свойства? Сколько еще существует в природе вирусов специализированных и универсальных?
На все эти вопросы лишь предстоит ответить.
С вирусами связано немало загадочного, неясного, а если быть точным до конца — еще не выясненного.
Признавая существование возбудителей инфекционных болезней, по размерам намного меньших, чем бактерии, ученые долго не могли прийти к единому мнению: какие они? Так, известный голландский микробиолог М. Бейеринк, к примеру, предполагал, что вирусы — необъяснимая загадка. Он дал им название Cоntagium vivum fluidum — живое жидкое заразное начало.
Другие исследователи пытались связать данные о вирусах с привычными для них представлениями о живом организме (клеточное строение, размножение путем деления с последующим ростом до размеров взрослой особи и т. д.). Не будем перечислять здесь другие предположения, высказанные на заре развития вирусологии. Все они — как наивные, так и наделенные долей предвидения — строились на одних лишь догадках, вслепую.
Много неясного и в современных гипотезах о происхождении вирусов. Так, одни исследователи считают, что вирусы — это потомки древних доклеточных форм жизни, застывшие, остановившиеся в своем развитии на определенном этапе. Разнообразие генетического вещества, говорят сторонники гипотезы, отражает ход эволюции этих существ. Природа как бы опробовала на вирусах все возможные варианты наследственного вещества, прежде чем остановиться окончательно на двухспиральной ДНК.
Вирусы — потомки бактерий или других одноклеточных организмов, по неизвестным причинам двинувшиеся в своем развитии вспять, деградировавшие, говорят другие ученые. Возможно, некогда их устройство было сложней, но со временем они многое утратили, и их нынешнее состояние, в том числе и разнообразие носителей генетической информации, лишь отражает разные уровни деградации, которых достигли различные их виды.
Наконец, существует гипотеза, согласно которой вирусы представляют собой составные части клеток живых существ, по неизвестной причине ставшие автономными системами. Процесс возникновения вирусов, согласно этой гипотезе, относится не только к глубокой древности, когда они уже, безусловно, существовали, но и к нашему времени. Иными словами, эта гипотеза признает возможность повсеместного, происходящего непрерывно образования вирусов клеточными элементами. Возможно ли такое, способны ли составные части клеток стать автономными, да еще и саморепродуцирующимися (способными к воспроизведению) системами?
Логика и парадоксы микромира
Устройство вирусов поражает своей чисто математической завершенностью, логикой симметрии. Возьмем, к примеру, наиболее просто организованный вирион (зрелый вирус) табачной мозаики.
Сотни белковых кристаллообразных структур уложены в виде тугой спирали. Сердцевина нити, образующей спираль, представляет собой своеобразную капсулу, где находится молекула нуклеиновой кислоты. В результате общий вид вириона — предельно лаконичный цилиндр, полая трубка.
А вот другая форма: двадцатигранник, икосаэдр, грани которого образованы треугольниками. Основной материал, из которого сложен икосаэдр, — те же белковые структуры. Внутри — полость, где покоится молекула нуклеиновой кислоты. Это вирион полиомиелита.
— Позвольте, — возражали многие ученые еще в недавнем прошлом, — да можно ли вообще после этого называть вирусы живыми существами? Может быть, это кристаллообразные вещества, наделенные болезнетворными свойствами?
— Либо, — говорили другие, — это пограничные формы между живым и неживым мирами.
Кто же прав? Скорей всего наиболее многочисленная группа исследователей, которая считает, что вирусы — представители живой природы, го есть не вещества, а существа. Правда, существа крайне своеобразные, ведущие сугубо паразитический образ жизни.
Вирус проникает в клетку
Вирусы, имеющие иное строение, проникают в клетку не столь затейливым путем. Притянутые к оболочке клетки и воздействующие на нее ферментами, они провоцируют втягивание внутрь того участка мембраны, на котором осели. Образуется своего рода капсула-вакуоль с вирусной частицей внутри. Вакуоль эта затем отрывается, и в ней, путешествующей внутри клетки, продолжают идти одновременно два процесса — вирусная частица с помощью своих ферментов разрушает окутывающие ее стенки капсулы, а ферменты клетки разрушают внешние оболочки вируса, освобождая, как это было и в случае с фагом Т2, нуклеиновую кислоту.
Итак, нуклеиновая кислота покинула белковую оболочку и исчезла, бесследно растворилась в клеточной среде. Что же дальше?
Мы еще не имеем возможности получить полный ответ на этот вопрос. До сих пор удалось установить характер лишь некоторых изменений, происходящих на этом этапе в различных частях клетки. И по этим отдельным штрихам мы воссоздаем, пытаемся представить себе полностью происходящее.
Формирование вирусов начинается, по-видимому, с подавления нормальных процессов обмена веществ в клетке. Установлено, в частности, что рибонуклеиновая кислота (РНК) вируса гриппа способна синтезировать на клеточных элементах — рибосомах, ведающих выработкой белка,— особое вещество, также белковой природы,— гистон, который, в свою очередь, связывается с ДНК клетки и прекращает синтез клеточной РНК. Некоторые другие вирусы, например, вирусы полиомиелита, не нуждаются в окольном пути, так как сами способны вмешаться в деятельность рибосом и прекратить синтез клеточных белков. Выявлены и другие механизмы подавления вирусами клеточного обмена, их вмешательства в жизнедеятельность клетки, но в конечном счете все сводится к одному: клеточные ресурсы перестают расходоваться на нужды самих клеток и поступают в распоряжение вирусной нуклеиновой кислоты.
Беззащитна ли клетка!
Цикл превращений, связанных с размножением вирусов, как правило, краток. В одних случаях проникновение вирусной нуклеиновой кислоты в клетку отделяет от появления вирионов 13—15 минут, в других — 40 минут. Вирусы одной из наиболее распространенных инфекций, гриппа, проходят этот путь примерно за 6—8 часов. И каждый раз около погибшей клетки оказываются десятки, а порой и сотни вирионов. Причем каждый из них, в свою очередь, готов к продолжению процесса размножения. Количество вирусной инфекции нарастает буквально лавинообразно.
Но так как главное действующее лицо — вирус остается за кадром (в обычный микроскоп он не виден), на экране только последствия его агрессии. Картина перед наблюдателем разворачивается впечатляющая. Вначале крайние клетки, первыми подвергшиеся нападению, начинают терять свойственные им округлые очертания. Постепенно истончаются их мембраны, клеточные элементы, клетка как бы взрывается. В этот момент, как мы знаем (но не видим этого), опустошенную оболочку покидают полчища вирионов, направляющихся к очередным своим жертвам. И через самое непродолжительное время точно так же изменяются, а затем лопаются соседние клетки, за ними другие, еще и еще.
. Колония клеточной культуры как бы охвачена пламенем. Вот она рассечена обезжизненными структурами на островки. Вот сжимаются и эти островки, уменьшаются в размерах, и. все кончено. Колония разрушена дотла.
Обладай вирусы такими же возможностями в естественных условиях, и человеку и любому другому живому существу пришлось бы плохо. Однако этого не происходит, ибо на страже — отработанные за миллионы лет защитные приспособления организма, ограничивающие могущество вирусов.
Безграничному расширению вирусной агрессии препятствуют прежде всего сами вирусы. Еще в 30-х годах ученые заметили, что размножение в клетке одного вируса нередко препятствует размножению в этой же клетке другого вируса.
Кстати, если говорить серьезно, одна из многочисленных гипотез, пытавшихся объяснить это явление, так и гласила: всему причиной конкуренция вирусов, борющихся за клеточные компоненты. Без малого три десятилетия понадобилось, чтобы раскрыть существо этого явления, получившего название интерференции. И, как оказалось, в данном случае инициатива принадлежала не вирусам, а самой клетке. На проникновение вируса (чему воспрепятствовать клетка, увы, не может) она отвечает немедленной выработкой особого белкового вещества — интерферона. Правда, интерферон не спасает уже пораженную клетку, но препятствует продвижению вирусной инфекции к другим клеткам организма. Иными словами, за первыми же вирионами, прорвавшимися в организм, возникает барьер интерфероновой защиты.
Антитела, появляющиеся позже, существуют несравненно дольше. Именно они и становятся основой стойкого иммунитета, благодаря которому многие инфекционные болезни не повторяются дважды в жизни одного индивидуума.
Медицина — в наступлении
Среди инфекционных заболеваний 80 процентов вирусных. Эта цифра — свидетельство победы человека над бактериальными инфекциями. Чума, холера, тиф, некогда безоговорочно первенствовавшие в медицинских статистических сводках, с приходом антибиотиков и сульфопрепаратов навсегда сдали свои позиции. Их место заняли болезни, вызываемые вирусами.
Как известно, и с этими недугами ведется успешная борьба. Побежден полиомиелит. Тягостным воспоминанием ушла в прошлое оспа. Широким фронтом идет наступление на корь: лишь за последнее пятилетие число перенесших заболевание корью снизилось в 5 раз; на повестке дня — полное искоренение этой инфекции на территории нашей страны.
Значительные усилия направляются на борьбу с гепатитом, гриппом, паротитом, вирусными респираторными заболеваниями, однако здесь решающие достижения еще впереди.
Наряду с этим ученые работают над созданием других эффективных лекарственных веществ, способных подавить вирусную инфекцию.
Работа эта начата. Во все концы нашей страны и за рубеж отправляются специальные экспедиции вирусологов. Уже получены чрезвычайно ценные данные о перемещениях вирусной гриппозной инфекции из Всемирного противогриппового центра, в деятельность которого вносит существенный вклад региональный противогриппозный центр СССР.
Бактериофаги – это вирусы, которые поражают только бактерий. В ходе инфекции они влияют на все процессы жизнедеятельности бактериальной клетки, фактически превращая ее в фабрику по производству вирусного потомства. В конце концов клетка разрушается, а вновь образованные вирусные частицы выходят наружу и могут заражать новые бактерии.
Несмотря на огромное число и разнообразие природных фагов, встречаемся мы с ними редко. Однако бывают ситуации, когда деятельность этих вирусов не остается незамеченной. Например, на предприятиях, где производят сыры, йогурты и другие молочно-кислые продукты, часто приходится сталкиваться с вирусной атакой на бактерии, сбраживающие молоко. В большинстве таких случаев фаговая инфекция распространяется молниеносно, и полезные бактерии гибнут, что приводит к значительным экономическим потерям (Neve et al., 1994).
Именно благодаря прикладным исследованиям в интересах молочной промышленности, направленным на получение устойчивых к бактериофагам штаммов молочно-кислых бактерий, был открыт ряд механизмов, с помощью которых бактерии избегают инфекции. Параллельно были изучены способы, с помощью которых вирусы, в свою очередь, преодолевают бактериальные системы защиты (Moineau et al., 1993).
Кто защищен – тот вооружен
На сегодня известно пять основных, весьма хитроумных механизмов защиты, которые бактерии выработали в непрестанной борьбе с вирусами: изменение рецептора на поверхности клетки; исключение суперинфекции; системы абортивной инфекции; системы рестрикции-модификации и, наконец, системы CRISPR-Cas.
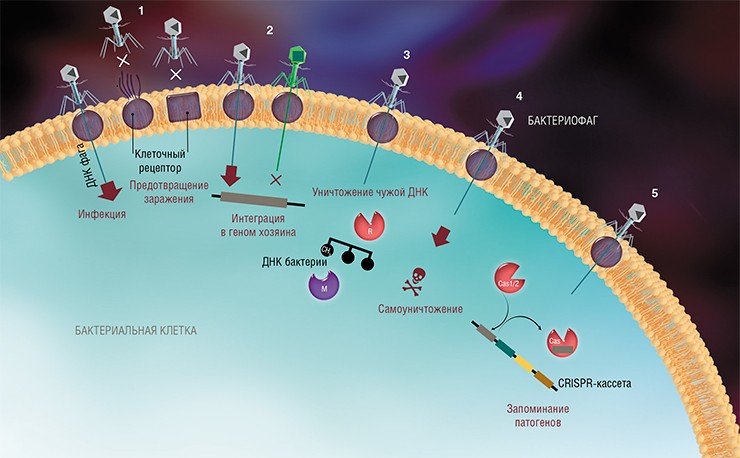
К средствам противовирусной защиты бактерий относятся и системы рестрикции-модификации, в которые входят гены, кодирующие два белка-фермента – рестриктазу и метилазу. Рестриктаза узнает определенные последовательности ДНК длиной 4—6 нуклеотидов и вносит в них двуцепочечные разрывы. Метилаза, напротив, ковалентно модифицирует эти последовательности, добавляя к отдельным нуклеотидным основаниям метильные группы, что предотвращает их узнавание рестриктазой.
Врага нужно знать в лицо
Системы CRISPR-Cas являются уникальным примером адаптивного иммунитета бактерий. При проникновении в клетку ДНК фага специальные белки Cas встраивают фрагменты вирусной ДНК длиной 25—40 нуклеотидов в определенный участок генома бактерии (Barrangou et al., 2007). Такие фрагменты называются спейсерами (от англ. spacer – промежуток), участок, где происходит встраивание, – CRISPR-кассета (от англ. Clustered Regularly Interspaced Short Palindromic Repeats), а сам процесс приобретения спейсеров – адаптацией.
Чтобы использовать спейсеры в борьбе с фаговой инфекцией, в клетке должен происходить еще один процесс, управляемый белками Cas, названный интерференцией. Суть его в том, что в ходе транскрипции CRISPR-кассеты образуется длинная молекула РНК, которая разрезается белками Cas на короткие фрагменты – защитные криспрРНК (крРНК), каждая из которых содержит один спейсер. Белки Cas вместе с молекулой крРНК образуют эффекторный комплекс, который сканирует всю ДНК клетки на наличие последовательностей, идентичных спейсеру (протоспейсеров). Найденные протоспейсеры расщепляются белками Cas (Westra et al., 2012; Jinek et al., 2012).
Системы CRISPR-Cas обнаружены у большинства прокариот – бактерий и архей. Хотя общий принцип действия всех известных систем CRISPR-Cas одинаков, механизмы их работы могут существенно отличаться в деталях. Наибольшие различия проявляются в строении и функционировании эффекторного комплекса, в связи с чем системы CRISPR-Cas делят на несколько типов. На сегодняшний день описаны шесть типов таких неродственных друг другу систем (Makarova et al., 2015; Shmakov et al., 2015).
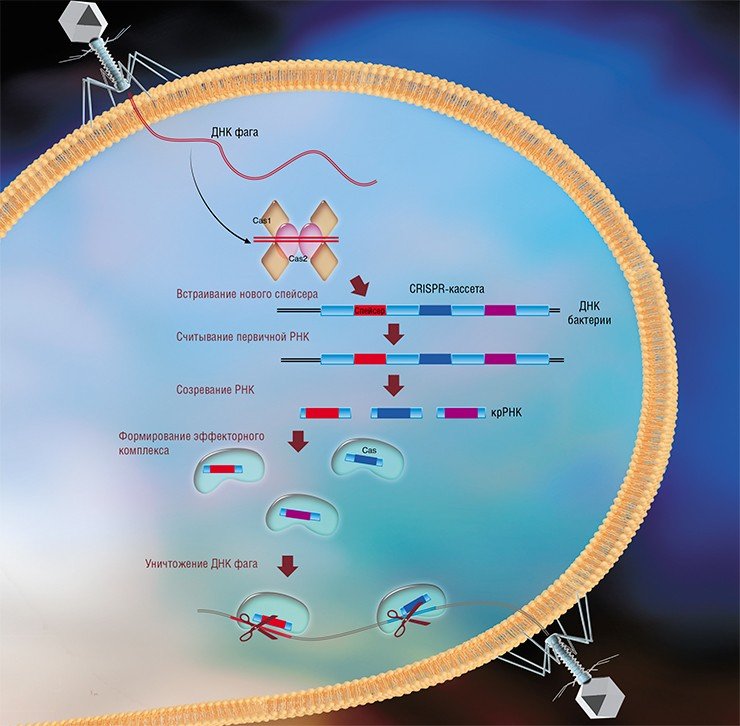
Наиболее изученной является система CRISPR-Cas I типа, которой обладает излюбленный объект молекулярно-биологических исследований – бактерия кишечная палочка (Esсherichia coli). Эффекторный комплекс в этой системе состоит из нескольких небольших белков Cas, каждый из которых отвечает за разные функции: разрезание длинной некодирующей CRISPR РНК, связывание коротких крРНК, поиск, а затем разрезание ДНК-мишени.
Гонка вооружений
Бактериофаги, как факторы среды, вызывают направленные изменения в геноме бактерий, которые наследуются и дают бактериям явное преимущество, спасая от повторных инфекций. Поэтому системы CRISPR-Cas можно считать примером ламарковской эволюции, при которой происходит наследование благоприобретенных признаков (Koonin et al., 2009)
Некоторые бактериофаги реагируют на наличие в бактериальной клетке систем CRISPR-Cas выработкой особых анти CRISPR-белков, способных связываться с белками Cas и блокировать их функции (Bondy-Denomy et al., 2015). Еще одно ухищрение — обмен участков генома вируса, на которые нацелена система CRISPR-Cas, на участки геномов родственных вирусов, отличающихся по составу нуклеотидной последовательности (Paez-Espino et al., 2015).
Благодаря постоянному совершенствованию биоинформатических алгоритмов поиска, а также включению в анализ все большего количества прокариотических геномов, открытие новых типов CRISPR-Cas систем является делом недалекого будущего. Предстоит также выяснить и детальные механизмы работы многих недавно открытых систем. Так, в статье, опубликованной в 2016 г. в журнале Science и посвященной анализу системы CRISPR-Cas VI типа, описан белок С2с2, образующий эффекторный комплекс с крРНК, который нацелен на деградацию не ДНК, а РНК (Abudayyeh et al., 2016). В будущем такое необычное свойство может быть использовано в медицине для регулирования активности генов путем изменения количества кодируемых ими РНК.
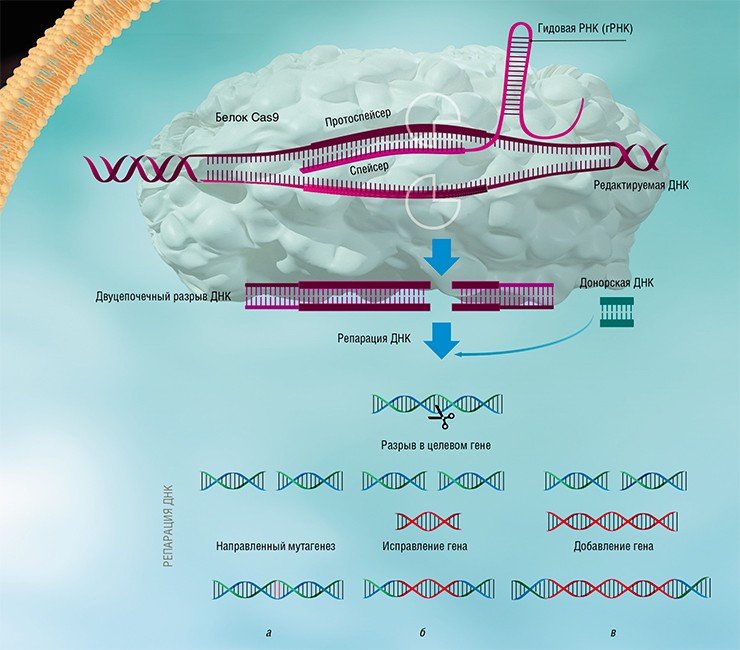
Изучение стратегий борьбы бактерий с бактериофагами, несмотря на свою кажущуюся фундаментальность и отвлеченность от задач практической медицины, принесло неоценимую пользу человечеству. Примерами этого могут служить методы молекулярного клонирования и редактирования геномов – направленного внесения или удаления мутаций и изменения уровня транскрипции определенных генов.
Благодаря быстрому развитию методов молекулярной биологии всего лишь через несколько лет после открытия механизма действия систем CRISPR-Cas была создана работающая технология геномного редактирования, способная бороться с болезнями, ранее считавшимися неизлечимыми. Доступность и простота этой технологии позволяют рассматривать ее как основу для медицины, ветеринарии, сельского хозяйства и биотехнологий будущего, которые будут базироваться на направленных и безопасных генных модификациях.
Нет никаких сомнений, что дальнейшее изучение взаимодействия бактерий и их вирусов может открыть перед нами такие возможности, о которых мы сейчас даже не подозреваем.
Abudayyeh O. O., Gootenberg J. S., Konermann S. et al. C 2c2 is a single-component programmable RNA-guided RNA-targeting CRISPR effector // Science. 2016. V. 353: aaf5573.
Barrangou R., Fremaux C., Deveau H. et al. CRISPR provides acquired resistance against viruses in prokaryotes // Science. 2007. V. 315. P. 1709–1712.
Bikard D., Marraffini L. A. Innate and adaptive immunity in bacteria: mechanisms of programmed genetic variation to fight bacteriophages // Curr. Opin. Immunol. 2012. V. 1 P. 15–20.
Bondy-Denomy J., Garcia B., Strum S. et al. Multiple mechanisms for CRISPR-Cas inhibition by anti-CRISPR proteins // Nature. 2015. V. 526. P. 136–139.
Calendar R., Abedon S. T. The Bacteriophages // 2nd Ed., Oxford University Press. 2006.
Datsenko K. A., Pougach K., Tikhonov A. et al. Molecular memory of prior infections activates the CRISPR/Cas adaptive bacterial immunity system // Nat. Commun. 2012. V. 3. P. 945
Jiang W., Marraffini L. A. CRISPR-Cas: New Tools for Genetic Manipulations from Bacterial Immunity Systems // Annu. Rev. Microbiol. 2015. V. 69. P. 209–28.
Jinek M., Chylinski K., Fonfara I., et al. A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity // Science. 2012. V. 337. P. 816–821.
Koonin E. V., Wolf Y. I. Is evolution Darwinian or/and Lamarckian? // Biol. Direct. 2009. V. 4. P. 42.
Lopez-Pascua L., Buckling A. Increasing productivity accelerates host-parasite coevolution // J. Evol. Biol. 2008. V. 3. P. 853–860.
Makarova K. S., Wolf Y. I., et al. An updated evolutionary classification of CRISPR-Cas systems // Nat. Rev. Microbiol. 2015. V. 11. P. 722–736.
Moineau, S., Pandian S., Klaenhammer T. R. Restriction/modification systems and restriction endonucleases are more effective on lactococcal bacteriophages that have emerged recently in the dairy industry // Appl. Envir. Microbiol. 1993. V. 59. P. 197–202.
Neve H., Kemper U., et al. Monitoring and characterization of lactococcal bacteriophage in a dairy plant // Kiel. Milckwirtsch. Forschungsber. 1994. V. 46. P. 167–178.
Nuñez J. K., Harrington L. B., et al. Foreign DNA capture during CRISPR-Cas adaptive immunity // Nature. 2015a. V. 527. P. 535–538.
Nuñez J. K., Kranzusch P. J., et al. Cas1-Cas2 complex formation mediates spacer acquisition during CRISPR-Cas adaptive immunity // Nat. Struct. Mol. Biol. 2014. V. 21. P. 528–534.
Nuñez J. K., Lee A. S., Engelman A., Doudna J. A. Integrase-mediated spacer acquisition during CRISPR-Cas adaptive immunity // Nature. 2015b. V. 519. P. 193–198.
Paez-Espino D., Sharon I., et al. CRISPR Immunity Drives Rapid Phage Genome Evolution in Streptococcus thermophilus // MBio. 2015. V. 6: e00262–15.
Shmakov S., Abudayyeh O. O., Makarova K. S., et al. Discovery and Functional Characterization of Diverse Class 2 CRISPR-Cas Systems. // Mol. Cell. 2015. V. 60. P. 385–397
Tan D., Svenningsen S. L., Middelboe M. Quorum sensing determines the choice of antiphage defense strategy in Vibrio anguillarum. // mBio 2015. V. 6: e00627–15.
Westra E. R., van Erp P. B., Künne T., et al. CRISPR immunity relies on the consecutive binding and degradation of negatively supercoiled invader DNA by Cascade and Cas3 // Mol. Cell. 2012. V. 46. P. 595–605.
Работа поддержана грантом РФФИ (№ 16-34-01176)
Ключевые слова
биология бактриофагов, экология бактерифагов, эволюция бактериофагов, адсорбционный аппарат бактериофагов, взаимодействия фаг — клетка, фаговая терапия, экология кишечника, высокоплотные микробные сообщества
Направления исследований
- Исследования структуры, механизмов функционирования и путей адаптации адсорбционных аппаратов бактериофагов
- Исследования экологии бактериофагов в природных симбиотических и свободноживущих высокоплотных бактериальных сообществах
- Исследования механизмов адсорбционной резистентности бактерий к фагам и механизмов ее преодоления вирусами
- Исследования биологического разнообразия фагов
- Исследования экологической структуры и взаимодействий бактерий в сообществах меромиктических озер
Основные методы исследования
Лаборатория проводит фундаментальные исследования в области экологии бактериофагов сочетая современные высокоразрешающие молекулярные методы типирования изолятов фагов и бактерий. В лаборатории осуществляется полный цикл от выделения и биологической характеристики бактериофагов, позволяющий вводить в практику новые модельные системы фаг-хозяин, пригодные для детальной расшифровки молекулярных механизмов экологически значимых взаимодействий, находить и описывать новые группы вирусов бактерий, исследовать разнообразие и динамику природных вирусных сообществ.
Методы фундаментальных исследований (анализ с фото- и флуориметрической детекцией):
- микробиологические и вирусологические методы
- методы генеомики и метагеномики
- методы генной и белковой инженерии и генетического анализа
- методы биохимии белков
- электронная и световая микроскопия
- методы структурной биологии (в колаборациях)
Краткая история лаборатории
Лаборатория создана решением ученого совета ИНМИ РАН в 2003 г. вместо лаборатории вирусологии, которая тем же решением ученого совета была выведена из состава института. И.о. заведующего лабораторией назначен к.б.н. А.В. Летаров.
Тематикой лаборатории с первых лет ее существования была расшифровка молекулярных механизмов, лежащих в основе экологической адаптации бактериофагов в природных микробных сообществах, разработка научных основ рациональной фаговой терапии.
Достижения
ОСНОВНЫЕ ДОСТИЖЕНИЯ
Работа лаборатории является логическим продолжением проектов, посвященных исследованию экологии бактериофагов в симбиотических микробных сообществах (см. Golomidova et al. 2007) и эволюции бактериофагов (Letarov et al. 2005, Comeau 2007). В рамках этой работы получены следующие результаты и достижения:
Исследовано влияние модификации О-антигена у E.coli 4s на характер чувствительности к бактериофагам, в результате чего продемонстрировано сильное влияние О-ацетилирования на эффективность неспецифической защиты клеток (Knirel et al. 2015).
В результате исследований бактериофагов, полученных в ходе проекта по экологии колифагов в кишечном микробном сообществе лошадей, изучены биологические свойства, определены и аннотированы полные геномные последовательности более 10 бактериофагов (Kulikov et al. 2012, Куликов с соавт. 2014, см. также GenBank KM979355.1 NC_019520.1 KM979354 еще несколько последовательностей готовятся к публикации).
Раскрыты молекулярные механизмы ранних стадий инфекции клеток Е.coli 4s N4- подобным фагом G7C (Kulikov et al. 2012). В частности, установлена пространственная структура одного из адгезинов этого вируса (PDB ID 4QNL ‘on hold’), установлена ферментативная активность этого белка (деацетилаза О-антигена ЛПС), Показано, что адгезины этого вируса имеют разветвленное строение при кооперативном действии компонентов (Prokhorov et al. 2014. VoM III, EMBO, abstracts; полная публикация в стадии подготовки).
Описан феномен формирования длительно персистирующих ассоциаций ряда вирулентных колифагов (G7C, 9G) с клетками и проведены исследования механизмов этого явления (Летарова с соавт. 2013, Kulikov et al. 2014, Letarova and Letarov 2013, 2012).
Осуществлено теоретическое обобщение имеющихся данных о роли бактериофагов в микробиоте тела животных и человека. Обзор (Letarov and Kulikov, 2009) явился первым таким обобщением за последние несколько десятилетий. Теоретические концепции и анализ новых данных были развиты в последующих работах Clookie et al. 2011, Letarov 2012). Приложение этих теоретических разработок в области фаговой терапии развито в работе Летаров с соавт. (2010). Нами был выполнен перевод на русский язык книги Э. Каттер и А. Сулкавелидзе ред, 2012 Бактериофаги. Биология и практическое применение. М. Научный мир.; (англ. издание 2005), которая вышла под научной редакцией А.В. Летарова. Русская версия была существенно дополнена с целью приведения в соответствие с наиболее современными достижениями области, в том числе включением двух новых врезок. Эта книга явилась первым фундаментальным изданием, посвященным биологии вирусов прокариот, вышедшим на русском языке с 1980-х гг.
В лаб. вирусов микроорганизмов ИНМИ РАН развита методология исследований экологии бактериофагов с учетом внутривидового (штаммового) разнообразия популяций бактерий – хозяев. Были существенно усовершенствованы ранее предложенные нами методы геномного ПЦР-фингерпринтинга энтеробактерий (Исаева с соавт. 2010). Приложение этой идеологии к исследованиям вагинального сообщества лактобацилл дало оценки внутривидового разнообразия индивидуальных популяций (Исаева с соавт. 2012, Исаева с соавт. 2013)
Выполнены исследования взаимодействий фагов и макроорганизма.
В настоящее время усилиями специалистов центра Коллекция формируется как депозитарий патогенных микроорганизмов. В коллекционном фонде содержатся:
 |
|
|  |
Целью работы коллекции является обеспечение правовой охраны штаммов микроорганизмов, а также гарантированное сохранение микроорганизмов в жизнеспособном состоянии и создании условий для длительного хранения с целью стандартизации исследований с различными микроорганизмами.
В задачи исследований коллекции входит:
|  | |
 |
| |
|  | |
 |
| |
|  | |
Сотрудники Коллекции организуют и проводят работы по подготовке ежегодной информации в рамках Конвенции о запрещении разработки, производства и накопления запасов бактериологического (биологического) и токсинного оружия (КБТО), а также участвуют в системе контроля трансграничного перемещения патогенных микроорганизмов.


Общую информацию о коллекции можно скачать здесь.
Коллекция включена в Государственный регистр информационных ресурсов 21.11.2012 № 1761203264
Адрес:
ул. Филимонова, д. 23
220114, г. Минск, Республика Беларусь
Читайте также:
