
Микоплазмы и микобактерии туберкулеза
Сайт СТУДОПЕДИЯ проводит ОПРОС! Прими участие :) - нам важно ваше мнение.
196.Экологический профиль большинства микобактерий:
1. Симбионты-комменсалы животных. 2. Симбионты-паразиты животных. 3. Симбионты-паразиты человека. 4. Сапрофиты.5. Облигатно-патогенные виды.
197.Возбудители микобактериальных антропонозов:
1. Mycobacterium bovis. 2. Mycobacterium leprae. 3. Mycobacterium avis/intarcellulare. 4. Mycobacterium tuberculosis. 5. Mycobacterium bovis, ssp. BCG.
198.Возбудитель микобактериального зооноза:
1. Mycobacterium bovis. 2. Mycobacterium leprae. 3. Mycobacterium kansasii. 4. Mycobacterium tuberculosis. 5. Mycobacterium bovis, ssp. BCG.
199.Возбудители туберкулеза человека:
1. Mycoplasma pneumoniae. 2. Mycobacterium avis/intracellulare.3. Mycobacterium tuberculosis. 4. Mycobacterium leprae.5. Mycobacterium bovis.
200.Признаки микобактерий туберкулеза,связанные с
особенностями их клеточной стенки:
1. Кислотоустойчивость. 2. Медленная скорость размножения.3. Резистентность к фагоцитам. 4. Устойчивость во внешней среде. 5. Высокая чувствительность к антибиотикам.
201.Факторы и механизмы,определяющие тинкториальные,эколого-физиологические и патогенетические особенности микобактерий туберкулеза:
1. Капсулоподобный экзополисахарид. 2. Эндоспоры.
3. Склонность к трансформации в L-формы. 4. Липиды клеточной стенки. 5. Отсутствие пептидогликана.
202.Липиды/гликолипиды клеточной стенки микобактерий
туберкулеза:
1. Миколовые кислоты. 2. Корд-фактор. 3. Туберкулин.
4. Микозиды. 5. Сульфатиды. 6. Пептидогликан.
203.Последовательность событий при развитии туберкулезного (M. tuberculosis)процесса:
1. Проникновение возбудителя в альвеолы. 2. Т-зависимая
активация макрофагов. 3. Размножение возбудителя в
альвеолярных макрофагах. 4. Формирование неспецифической (доиммунной) гранулемы. 5. Индукция Т-клеточного иммунитета. 6. Проникновение возбудителя в регионарные лимфатические узлы. 7. Формирование специфической (постиммунной гранулемы).
204.В инициации инфекционного процесса участвуют следующие факторы и механизмы микобактерий туберкулеза:
1. Внутриэпителиальная инвазия. 2. Пиогенная инвазия
субэпителиальных тканей. 3. Внутримакрофагальная инвазия.4. Внутрисосудистая инвазия (бактериемия). 5. Деструктивные ферменты и токсины.
205.Туберкулин:
1. Комплекс липидов клеточной стенки туберкулезной палочки. 2. Комплекс туберкулопротеинов (белковых дериватов). 3. Обладает прямой болезнетворностью. 4. Обладает иммунологически опосредованной болезнетворностью. 5. Используется для вакцинопрофилактики туберкулеза.6. Используется в аллергодиагностике туберкулезной инфекции.
206.Положения,справедливые для белковых антигенов
туберкулезной палочки (туберкулопротеинов):
1. Обладают прямой болезнетворнстью. 2. Определяют
культуральные и тинкториальные особенности микобактерий.3. Проявляют имунологически зависимую болезнетворность.4. Участвуют в реализации протективного иммунитета.5. Используются в аллергодиагностике. 6. Обладают иммуноадъювантным эффектом.
207.Положения,справедливые для липидов/гликолипидов
клеточной стенки микобактерий туберкулеза:
1. Обладают антимакрофагальной активностью. 2. Участвуют в индукции Т-клеточного иммунитета (иммуноадъювантный эффект). 3. Действующее начало туберкулина. 4. Проявляют иммунологически зависимую болезнетворность. 5. Определяют культуральные и тинкториальные особенности микобактерий.
208.Главная функция,реализуемая макрофагами в зоне
неспецифической (доиммуной)гранулемы при уберкулезной
инфекции:
1. Уничтожение микобактерий. 2. Возбуждение реакций
клеточного (Т- зависимого) иммунитета. 3. Индукция реакций гуморального иммунитета. 4. Функциональная кооперация с Т- эффекторами. 5. Предупреждение микробной инвазии.
209.Положения,справедливые для специфической
(постиммунной)туберкулезной гранулемы:
1. Возникает на фоне аллергии замедленного типа к
туберкулопротеинам. 2. Зона для функциональной кооперации между макрофагами и Т-эффекторами. 3. Основа для деструктивных реакций. 4. Основа для саногенеза. 5. Обычно завершается бессимптомной персистенцией возбудителя.
210.Решаюшую роль в интоксикации при туберкулезе играют следующие факторы и механизмы:
1. Микобактериальные токсины. 2. Туберкулопротеины.
3. Антигенная стимуляция Т-лимфоцитов. 4. Т-зависимая
активация макрофагов. 5. Цитокины иммунокомпетентных
211.Деструктивные процессы при туберкулезе определяются следующими механизмами:
1. Прямое действие микобактериальных токсинов.
2. Иммунологически опосредованные эффекты
микобактериальных антигенов. 3. Цитокин-зависимая активация макрофагов.4. Аллергия к туберкулопротеинам.
5. Аллергия к липидам клеточной стенки микобактерий.
212.Наиболее вероятные исходы специфической (постиммунной)гранулемы:
1. Заживление (фиброз, кальцификация). 2. Полное уничтожение возбудителя. 3. Формирование латентной (персистентной) инфекции. 4. Генерализация процесса. 5. Формирование иммунитета к экзогенному реинфицированию.
213.Положения,справедливые для противотуберкулезного
иммунитета:
1. Нестерильный иммунитет. 2. Антитоксический иммунитет.2. Антибактериальный иммунитет. 4. Гуморальный иммунитет.5. Клеточный иммунитет.
214.Положения,справедливые для первичного туберкулеза:
1.Результат экзогенного инфицирования. 2. Следствие
215.Положения,справедливые для вторичного туберкулеза:
1. Реактивационный туберкулез. 2. Может быть следствием
экзогенного реинфицирования. 3. Хроническая патология легких (типичное течение). 4. Развивается на фоне аллергии к туберкулопротеинам. 5. Результат прямого действия
216.В аллергодиагностике туберкулезной инфекции используют препарат на основе следующих продуктов микобактерий:
1. Липиды/гликолипиды. 2. Пептидогликан. 3. Протеины.
4. Нуклеиновые кислоты. 5. Полисахариды.
217.Диагностическая значимость бактериоскопии (мокроты)при туберкулезе основана на следующем признаке возбудителя:
1. Морфологические особенности. 2. Образование эндоспор.
3. Подвижность. 4. Тинкториальное своеобразие.
5. Образование внутриклеточных включений.
218.Метод,на котором основана классическая экспресс-
диагностика туберкулеза:
1. Бактериоскопия. 2. Выявление микобактериальных антигенов.3. Ускоренные культуральные методы 4. Выявление специфических метаболитов. 5. Обнаружение специфических фрагментов ДНК (ПЦР).
219.Наиболее очевидные метаболические процессы,которые могут служить мишенью для специфических
противотуберкулезных антибиотиков:
1. Синтез пептидогликана. 2. Синтез белка. 3. Синтез
нуклеиновых кислот. 4. Синтез миколовых кислот.
5. Организация межмолекулярных контактов в клеточной стенке
.220.Положения,справедливые для туберкулезной вакцины (BCG):
1. Живая аттенуированная вакцина. 2. Анатоксин. 3. Убитая вакцина.4. ДНК-вакцина. 5. Конъюгированная вакцина.
2.10.РИККЕТСИИ
221.Признаки,отражающие атипичность риккетсий:
1. Отсутствие рибосом. 2."Энергетические паразиты".
3. Факультативные внутриклеточные паразиты. 4. Отсутствие пептидогликана. 5. "Мембранные паразиты".
222.Позиции,справедливые для риккетсий:
1. Зависимость от энергетического метаболизма клеток.
2. Размножение путем деления. 3. Дизъюнктивный способ
репродукции. 4. Размножение внутри фагосом. 5. Размножение в цитоплазме и/или ядре.
223.Признаки,характерные для экологии риккетсий:
1.Симбионты кровососущих членистоногих. 2. Возможность
трансовариальной передачи у кровососущих насекомых.
3. Персистенция в организме животных.
4. Почвенный резервуар (сапрофитическая фаза).
5. Циркуляция в системе "животные--членистоногие".
224.Возможная позиция членистоногих в экологии риккетсий:
1.Естественный резервуар инфекции. 2. Вектор (переносчики) инфекции. 3. Поддержка циркуляции риккетсий среди животных.4. Стабилизация природных очагов риккетсиозной инфекции.5. Участие в эпидемическом процессе.
225.Механизмы,определяющие болезнетворность риккетсий:
1. Размножение в макрофагах региональных лимфатических узлов. 2. Риккетсемия. 3. Паразитирование внутри эпителиальных клеток. 4. Паразитирование внутри
эндотелиальных клеток. 5. Прямое поражение клеток нервной ткани. 6. Иммунологически зависимое поражение тканей.
226.Наиболее приемлемый для практики метод лабораторной диагностики риккетсиозов:
1. Выделение и идентификация чистой культуры. 2. Выявление риккетсиозных антигенов в пораженных тканях. 3. Выявление специфических антител (серодиагностика). 4. Выявление генетических маркеров (полимеразная цепная реакция). 5. Биопробы на животных.
227.Риккетсии,для которых человек является природным
(естественным)резервуаром:
1. Rickettsia typhi. 2. Rickettsia prowazekii. 3. Rickettsia rickettsii. 4. Orientia tsutsugamushi. 5. Rickettsia sibirica.
228.Эпидемиологическая характерстика большинства
риккетсиозов,исходящая из экологии риккетсий:
1. Зооноз. 2. Антропоноз. 3. Трансмиссивный механизм передачи. 4. Природная очаговость. 5. Сапроноз.
229.Риккетсиозный антропоноз:
1. Лихорадка цуцугамуши. 2. Пятнистые лихорадки.
3. Эпидемический сыпной тиф. 4. Эндемичный сыпной тиф. 5. Лихорадка Ку.
230.Позиция человека в экологии риккетсий (за исключением R.prowazekii):
3. Обязательный участник жизненного цикла риккетсий.
4. Случайный хозяин. 5. Вектор инфекции.
231.Позиция вшей в экологии риккетсий эпидемического сыпного
тифа:
1.Обязательный участник жизненного цикла риккетсий.2. Природный резервуар инфекции. 3. Биологический вектор инфекции. 4. Механический вектор инфекции. 5. Естественный хозяин.
232.Существование риккетсий эпидемического сыпного тифа в природе поддерживается следующими факторами:
1. Социальное неблагополучие. 2. Персистенция возбудителя в организме переболевших людей. 3. Реактивация эдогенной инфекции. 4. Возможность внечеловеческого резервуара инфекции. 5. Трансовариальная передача возбудителя у вшей-переносчиков.
233.Экологическая система,поддерживающая существование R.prowazekiiв природе:
1. Грызуны -- клещи. 2. Крысы -- блохи. 3. Человек -- вши.
4. Человек -- вши -- животные. 5. Животные -- клещи -- человек.
234.Основная категория клеток,поражаемых при риккетсиозах:
1. Эпителиоциты. 2. Эритроциты. 3. Нейтрофилы.
4. Эндотелиоциты. 5. В-лимфоциты. 6. Т-лимфоциты.
235.Клиническое проявление реактивации персистентной R.prowazekii-инфекции:
1. Ку-лихорадка. 2. Болезнь Цинссера-Брилля. 3. Йерсинеоз.
4. Пситтакоз. 5. Трахома.
236.Экологическая система,поддерживающая существование
большинства риккетсий в природе:
1. Грызуны (мелкие млекопитающиеся) -- клещи. 2. Крысы --блохи. 3. Человек -- вши. 4. Человек -- вши -- животные.
5. Животные -- клещи -- человек.
237.Возбудитель эпидемического сыпного тифа:
1. Rickettsia prowazekii. 2. Rickettsia rickettsii. 3. R. sibirica.
4. Rickettsia (Orientia) tsutsugamushi. 5. Rickettsia typhi. 6. Coxiella burnetii.
238.Возбудитель эндемичного сыпного тифа:
1. Rickettsia prowazekii. 2. Rickettsia rickettsii. 3. R. sibirica.
4. Rickettsia Orientia) tsutsugamushi. 5. Rickettsia typhi. 6. Coxiella burnetii.
239.Риккетсии группы пятнистых лихорадок:
1. Rickettsia prowazekii. 2. Rickettsia rickettsii. 3. R. sibirica.
4. Rickettsia (Orientia) tsutsugamushi. 5. Rickettsia typhi.
240.Риккетсии группы сыпных тифов:
1. Rickettsia prowazekii. 2. Rickettsia rickettsii. 3. R. sibirica.
4. Rickettsia (Orientia) tsutsugamushi. 5. Rickettsia typhi.
241.Болезнь Цинссера-Брилля:
1.Эндогенная инфекция. 2. Реактивация персистентной инфекции (Rickettsia prowazekii). 3. Повторная экзогенная инфекция (Rickettsia prowazekii). 4. Эпидемиологически опасная инфекция. 5. Вариант эпидемического тифа.
2.10.ХЛАМИДИИ
242.Признаки,отражающие "атипичность"хламидий:
1. Неспособность к росту на искусственных питательных средах.2. Холестеролзависимость (мембранный паразитизм).
3. Дизъюнктивный способ репродукции. 4. Фильтруемость.
5. Отсутствие пептидогликана.
243.Положения,отражающие своеобразие репродукции хламидий:
1.Дизъюнктивный способ размножения. 2. Чередование фаз
элементарных и ретикулярных тел. 3. Синтез белка на рибосомах клетки-хозяина. 4. Размножение внутри эндосом (фагосом).5. Зависимость от энергетического метаболизма клетки-хозяина.
244.Для элементарных тел хламидий справедливы следующие положения:
1. Метаболическая инертность. 2. Фильтруемость.
3. Устойчивость во внешней среде. 4. Деление.
5. Трансформация в ретикулярные тела. 6. Инфекционность
(инфекционное начало хламидий).
245.Положения,справедливые для ретикулярных тел хламидий:
1. Репродуктивная (вегетативная) форма хламидий
2. Метаболическая инертность. 3. Способность к делению.
4. Трансформация в элементарные тела. 5. Устойчивость во
246.Причины относительной резистентности хламидий к
антибиотикам:
1. Отсутствие пептидогликана. 2. Внутриклеточный паразитизм.3.Метаболическая инертность элементарных тел.4. Метаболическая инертность ретикулярных тел. 5. Отсутствие молекулярных мишеней для ингибиторов белкового синтеза.
247.Хламидии --возбудители зоонозной инфекции:
1.Chlamydia trachomatis, серотипы A-C. 2. Chlamydia trachomatis, серотипы D-K. 3. Chlamydia trachomatis, серотипы L1-L3. 4. Chlamydia psittaci. 5. Chlamydia pneumoniae.
248.Хламидийный зооноз:
1. Трахома. 2. Генитальный хламидиоз. 3. Венерическая
лимфогранулема. 4. Пситтакоз/орнитоз. 5. Респираторный
хламидиоз (Ch. pneumoniae).
249.Хламидии,для которых человек служит естественным
хозяином (природным резервуаром):
1.Chlamydia trachomatis, серотипы A-C. 2. Chlamydia trachomatis, серотипы D-K. 3. Chlamydia trachomatis, серотипы L1-L3.4. Chlamydia psittaci. 5. Chlamydia pneumoniae.
.250.Высокая инвазивность характерна для следующих хламидий:
1. Chlamydia trachomatis, серотипы A-C. 2. Chlamydia trachomatis, серотипы D-K. 3. Chlamydia trachomatis, серотипы L1-L3.4. Chlamydia psittaci. 5. Chlamydia pneumoniae.
251.Патогенетические варианты Chlamdia trachomatis:
1. "Глазные" хламидии. 2. "Генитальные" хламидии.
3. "Респираторные" хламидии. 4. "Зоонозные" хламидии
5. Возбудители венерической лимфогранулемы.
252.Наиболее реальные методы лабораторной диагностики
генитального хламидиоза и носительства генитальных хламидий:
1. Выделение и идентификация чистой культуры.
2. Выявление антихламидийных антител. 3. Выявление
эпителиальных клеток с включениями (микроколониями
хламидий). 4. Выявление хламидийных антигенов
(иммуноферментный анализ). 5. Выявление фрагментов
хламидийной ДНК (полимеразная цепная реакция).
2.11.МИКОПЛАЗМЫ
253. "Атипичность"микоплазм подразумевает:
1. Облигатность внутриклеточного паразитизма.
2. Образование уникальных токсинов. 3. Облигатную
анаэробность. 4. Особенности строения бактериальных мембран.5. Отсутствие пептидогликана.
254.Позиции,справедливые для микоплазм:
1. Дизъюнктивный способ репликации. 2. Плеоморфизм.
3. Отсутствие пептидогликана. 4. Энергетические паразиты.5. Мембранные паразиты.
255.Позиции,справедливые для микоплазм:
1. Наличие многослойной липидной мембраны.
1. Фильтруемость. 3. Облигатность внутриклеточного
паразитизма. 4. Наличие ригидной клеточной стенки.
256.Термин "молликуты" (класс Mollicutes)отражает:
1. Структурное своеобразие клеточной оболочки.
2. Облигатность внутриклеточного паразитизма.
3. Ауксотрофность. 4. Особенности энергетического метаболизма.5. Патогенетическую значимость микоплазм.
257.Название микоплазм на уровне класса:
1. Риккетсии. 2. Хламидии. 3. Молликуты. 4. Прокариоты.
258.Родовые таксоны "медицинских"молликут:
1. Mycoplasma. 2. Chlamydia. 3. Haemophilus. 4. Ureaplasma.
5. Mycobacteria. 6. Helicobacter. 7. Neisseria. 8. Bordetella.
259.Антибиотики,не действующие на микоплазмы:
1. Ингибиторы цитоплазматической мембраны. 2. Бета-
лактамные антибиотики. 3. Ингибиторы синтеза пептидогликана.4. Ингибиторы синтеза белка. 5. Ингибиторы синтеза нуклеиновых кислот.
260.Признаки,по которым микоплазмы принципиально
отличаются от L-форм бактерий:
1. Отсутствие генов, детерминирующих синтез пептидогликана.2. Дефицит генов, детерминирующих синтез рибосомальных РНК.3. Нарушение синтеза пептидогликана. 4. Дефекты энергетического метаболизма. 5. Морфологическая нестабильность (дисбаланс клеточного роста).
261.Микоплазмы --первичные патогены человека:
1. Mycoplasma hominis. 2. Ureaplasma urealyticum. 3. Mycoplasma oralis. 4. Mycoplasma pneumoniae. 5. Chlamydia trachomatis.
262.Условно-патогенные микоплазмы --возбудители
урогенитальных микоплазмозов:
1. Mycoplasma hominis. 2. Ureaplasma urealyticum. 3. Mycoplasma oralis. 4. Mycoplasma pneumoniae. 5. Chlamydia trachomatis.
263.Микоплазмы,принимающие участие в образовании камней внутри мочевыводящей системы:
1. Mycoplasma hominis. 2. Ureaplasma urealyticum. 3. Mycoplasma oralis. 4. Mycoplasma pneumoniae. 5. Chlamydia trachomatis.
264.В реализации патогенетического потенциала микоплазм задействованы следующие факторы и механизмы:
1. Мембранный паразитизм. 2. Оксиданты. 3. Высокая
инвазивность. 4. Иммунологически опосредованная
агрессивность. 5. Прямое повреждение клеточных мембран.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Родовое название Mycobacterium (грибовидные бактерии) предложено Леманом и Нойманом в 1896 г., обративших внимание на плесневидную пленку, которая образуется при росте туберкулезной палочки на поверхности жидких сред (аэробы!). Вскоре коллекция микобактерий была дополнена видами, изолированными из внешней среды. Оказалось, что именно свободно живущие сапрофиты составляют большинство этой своеобразной группы прокариот, а истинно паразитические микобактерии представлены всего несколькими видами, патогенными для человека и животных. Впрочем, известную опасность представляют и некоторые микобактерии-сапрофиты (рис. 1). Они вызывают оппортунистические инфекции (микобактериозы) у иммунокомпрометированных лиц, а при заражении в раннем детстве могут извращать реактивность организма к возбудителям туберкулеза.
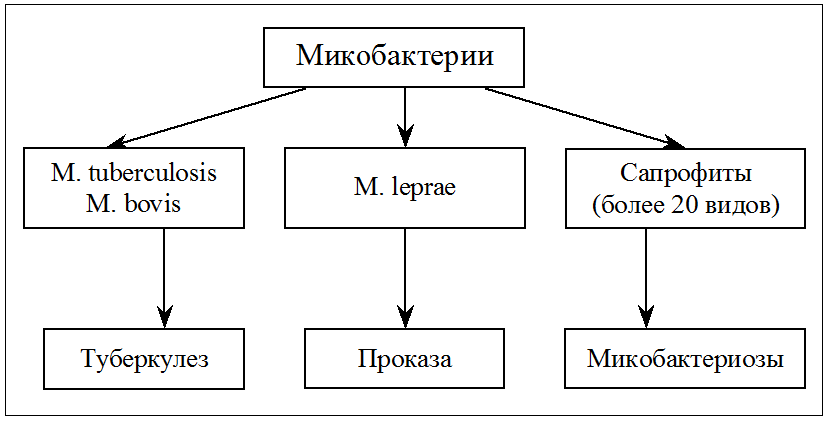
Рис. 1. Микобактерии, патогенные для человека
Известно более 60 видов микобактерий. Они объединены в род Mycobacterium семейства Mycobacteriaceae, которое относится к порядку Actinomycetales. В этом есть логика. Подобно актиномицетам, микобактерии образуют ветвящиеся клетки (особенно in vitro). Но сходство имеет и более глубокие корни, по крайней мере с соседями по порядку — нокардиями и коринебактериями. И те и другие содержат липиды, которые напоминают миколовые кислоты, хотя и уступают микобактериям по сложности строения этих уникальных компонентов клеточной стенки.
Микобактерии туберкулеза представляют собой тонкие, прямые или слегка изогнутые палочки, длиной 1—4 мкм и около 0,3 мкм в ширину (рис. 2). Они неподвижны, не образуют спор и капсулы, если не считать микозидной оболочки, которую иногда сравнивают с микрокапсулой. Они плохо окрашиваются по Граму, но, восприняв окраску, не обесцвечиваются этанолом, поэтому их считают грамположительными, хотя правильнее не относить ни к тем, ни к другим. С трудностями окраски туберкулезной палочки впервые столкнулся Кох, сумевший разглядеть их в мазках из мокроты лишь после 24—30-часовой (!) инкубации в щелочном растворе метиленовой синьки.
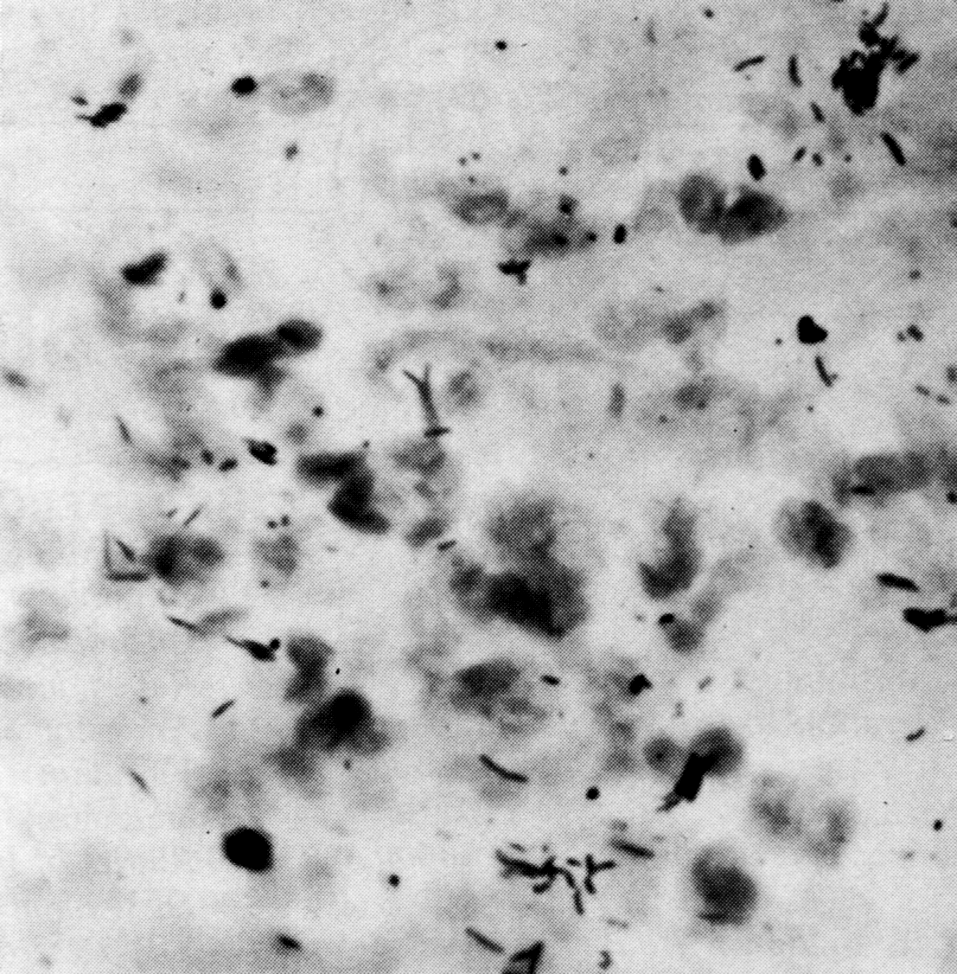
Рис. 2. M. tuberсulosis. Мазок из мокроты больного туберкулезом легких. Окраска по Цилю—Нильсену, х1050
Избирательная окраска основана на так называемой кислотоустойчивости микобактерий и включает два основных этапа. Сначала повышают проницаемость клеточной стенки для насыщенного раствора красителя (расплавление восковидного слоя путем прогревания или протравливания детергенами), а затем окрашенный препарат промывают разведенными кислотами. Обесцвечивая большинство бактерий, это сохраняет окраску микобактериальных клеток. По классической методике Циля—Нильсена на фиксированный мазок наливают карболовый фуксин, нагревают до отхождения паров, промывают закисленным этанолом (3% НСl в 85% растворе этанола) и докрашивают метиленовой синькой. Микобактерии удерживают фуксин и выглядят как красные палочки на голубом фоне; остальные бактерии теряют фуксин и окрашиваются в синий цвет. Этанол здесь не обязателен, но делает фон более чистым. Устойчивость к обесцвечиванию (она связана с образованием прочных комплексов между красителем и миколовыми кислотами клеточной стенки) проявляется не только в отношении кислот, но также щелочей и спиртов. Поэтому с равной справедливостью микобактерии можно называть также щелоче- и спиртоустойчивыми.
Морфологические и тинкториальные признаки микобактерий имеют много исключений, породивших немало споров о природе возбудителя. Туберкулезная палочка может терять кислотоустойчивость и хорошо окрашиваться по Граму, превращаться в филаментозные и даже мицелиоподобные формы, распадающиеся на палочки и кокки (гранулы/зерна Муха). Некоторые из них настолько малы, что проходят через бактериальные фильтры (фильтрующиеся формы). Теперь ясно: речь идет о фенотипической изменчивости, которая отражает экологическую пластичность возбудителя и его способность выживать в неблагоприятных условиях. Отсюда не удивительно, что плеоморфизм туберкулезной палочки сильнее проявляется в инвитровых культурах, т.е. в неестественной среде обитания.
Туберкулезные микобактерии — строгие аэробы и мезофилы, т.е. растут в диапазоне 30—42° С, лучше всего при 37°С. Размножение происходит очень медленно: период генерации составляет 14—16 ч (типичные бактерии делятся каждые 15 мин). Поэтому для получения обильного роста требуется не менее 4—6 нед, хотя миниатюрные колонии могут появиться через 7—10 дней. Туберкулезная палочка принадлежит к числу наиболее вяло реплицирующихся микобактерий. Большинство сапрофитических видов размножаются быстрее, их рост заметно отстает от других прокариот и хорошо заметен не ранее чем через 5—7 дней. Одной из причин отсроченного размножения микобактерий является высокая гидрофобность, связанная с обилием липидов в клеточной стенке. Это затрудняет поступление в бактерии питательных веществ, снижая их метаболическую активность.
Выделение первичных культур (т.е. непосредственно от больного) проводят на специальных средах, самые сложные из которых содержат яйца, картофельную муку и глицерин. Для подавления сопутствующей микрофлоры добавляют малахитовый зеленый или генцианвиолет. При субкультивировании туберкулезная палочка становится менее прихотливой и растет на обычных средах с добавкой глицерина.
В жидких средах рост происходит на поверхности (аэроб!). Нежная сухая пленка со временем утолщается, становится бугристо-морщинистой и обретает желтоватый оттенок, часто сравниваемый с цветом слоновой кости. Бульон остается прозрачным и добиться диффузного роста удается только в присутствии детергентов, например твина-80. В микроколониях (они образуются на ранних сроках и заметны только под микроскопом) формируются структуры, напоминающие жгуты — признак, который связывают с так называемым корд-фактором М. tuberculosis (рис. 3).

Рис. 3. Образование жгутов в микрокультуре М. tuberculosis
Морфотинкториальная изменчивость, о которой говорилось выше, весьма характерна для туберкулезной палочки, но это — фенотипические варианты, которые не переходят в генетически закрепленные биовары. Попытки выявить устойчивые серотипы М. tuberculosis и М. bovis не увенчались успехом, прежде всего из-за спонтанной агглютинации клеток, которая является следствием их высокой гидрофобности (см. ниже). Более удачным оказался опыт фаготипирования, но и он не закрепился в лабораторной практике.
В современных исследованиях много внимания уделяется дифференцировке (клонированию) микобактериальных штаммов по генетическим маркерам, прежде всего по особенностям хромосомного профиля ДНК. Генотипирование обычно проводится на основе вставочных генов — инсерционных последовательностей (англ. insertion sequences — IS). Они отличаются структурным полиморфизмом, позволяя классифицировать штаммы по степени генетического родства. Чаще используется анализ последовательности IS6110. Применяются и другие, дополнительные методы, основанные на особенностях генетического аппарата М. tuberculosis. К их числу относится сполиготипирование (от англ. spacer oligotyping), сравнение по числу прямых повторов и полиморфной G-С-обогащенной повторяющейся последовательности.
Полная идентичность возможна лишь внутри одного клона бактерий. Отсюда сравнение с отпечатками пальцев — англ. DNA fingerprinting. Каждое очередное поколение (т.е. каждый новый клон) несет хотя бы небольшие генетические различия. Накапливаясь со временем, они ведут к формированию клоногрупп, объединяемых в семейства. Тенденцию к распространению имеют клональные кластеры, еще не достигшие статуса группы. Наиболее изученным и распространенным (в том числе в России) является W-Beijing-семейство штаммов микобактерий туберкулеза. Оно включает более десятка клоногрупп — генетических ответвлений, эволюционировавших по IS6110 в различных географических зонах. Факторы, содействующие селекции штаммов W-Beijing, не известны. Возможно, это связано с повышенной контагиозностью, устойчивостью во внешней среде, резистентностью к антибиотикам. Именно первый W-штамм (акроним выбран произвольно), изолированный в 1990 г. в США, открыл тревожную эру лекарственной полирезистентности микобактерий туберкулеза. Есть мнение, что W-Beijing-штаммы более удачливы в преодолении барьера, создаваемого BCG-вакцинацией. Впрочем, каждая из перечисленных позиций встречает возражения.
Большую озабоченность вызывает эволюция лекарственной устойчивости на основе генетически закрепленных мутаций М. tuberculosis. Распространение резистентных биоваров может обезоружить в борьбе с туберкулезом или, по крайней мере, заметно снизить ее эффективность. Не случайно в системе лабораторий США, работающих с возбудителями особо опасных инфекций, организован центр, единственная задача которого — изучение штаммов туберкулезной палочки с множественной устойчивостью к антибиотикам.
Разгадка своеобразия микобактерий связана с необычностью их поверхностных структур. В клеточной стенке, устроенной сложнее, чем у других бактерий, преобладают липиды (более 60% сухой массы), в том числе специфичные для микобактерий. Именно они определяют нестандартность тинкториальных, физиологических и экологических свойств туберкулезной палочки (см. таблицу). Разнообразие микобактериальных липидов, в которых с трудом ориентируются даже искушенные биохимики, заставляет пользоваться такими собирательными понятиями, как миколовые кислоты, микозиды, сульфолипиды, корд-факторы и пр.
| Признак | Причина |
| Тинкториальные свойства: | |
| неокрашиваемость обычными способами | Слабая проницаемость клеточной стенки |
| кислото-, щелоче-, спиртоустойчивость | Связывание красителей миколовыми кислотами |
| Медленное размножение | Низкая скорость внутриклеточной диффузии питательных веществ |
| Культуральные свойства (сухие, морщинистые колонии), спонтанная агглютинация бактериальных клеток | Гидрофобность клеток |
| Устойчивость во внешней среде | Защита от высыхания |
| Устойчивость к дезинфектантам | Медленное проникновение антисептиков в клетку |
| Взаимоотношения с макрофагами: | |
| поглощение | Гидрофобность клеток |
| внутриклеточное выживание | Блокада образования фаголизосом, нейтрализация антимикробных факторов, ускользание в цитоплазму (повреждение фагосомальных мембран) |
| Цитотоксичность | Повреждение митохондриальной мембраны |
| Особенности иммунитета, иммунопатогенез | Иммуноадъювантная активность, CD1-зависимое представление антигенов (гликолипидов) |
Читайте также:
