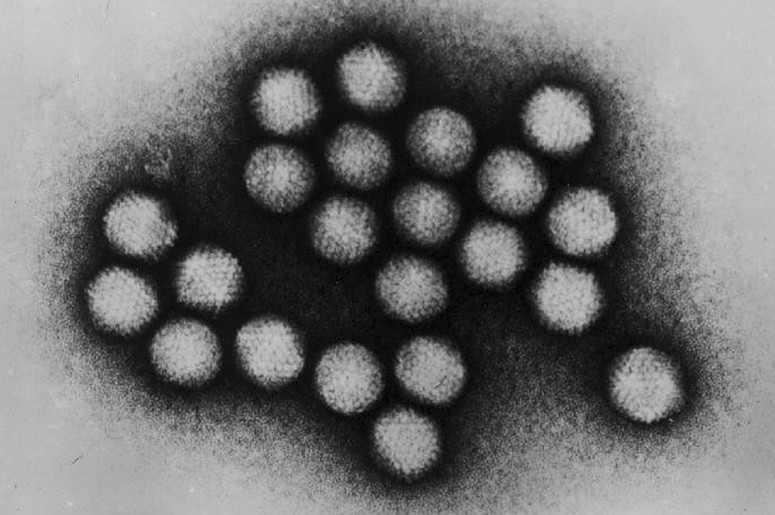
Вирусный капсид состоит из молекул
Вирус устроен предельно просто: молекула нуклеиновой кислоты в белковом чехле. Больше у него нет ничего, даже цвета. Но когда электронный микроскоп слой за слоем ощупает вирус, а умная компьютерная программа раскрасит его в яркие цвета, нашим глазам предстают такие композиции, которые сделали бы честь любой галерее абстрактного искусства.


1. Низкая электронная плотность
2. Высокая электронная плотность
3–5. Белковые оболочки
Лихорадка леса Cемлики
Заболевание эндемично для отдельных районов тропической Африки. Резервуаром вируса являются приматы, переносчиками — комары родов Aedes и Mansonia


1. Белки капсида
2. Вирусная ДНК
3–4. Белки портала
5. Белки хвоста
В качестве антибактериальной терапии острых кишечных инфекций, вызываемых сальмонеллами, часто используются бактериофаги. Они размножаются внутри бактерий и вызывают их разрушение (лизис)


1. Гликопротеиды внешней оболочки
2. Липиды
3. Белки капсида
Вирус циркулирует между птицами и комарами. Болеют в основном сельские жители. Поражения суставов в форме артрита могут сохраняться месяцами и даже годами
Вирус Синдбис получил имя по названию египетской деревни, где его впервые выделили. Однако такие же или почти такие же вирусы вызывают лихорадку, сыпь и боли в суставах у жителей Финляндии (болезнь Погоста), Швеции (болезнь Окельбо) и России (карельская лихорадка). Этот вирус устроен еще сложнее, чем его близкий родич SFV: внешняя оболочка состоит из гликопротеидов — соединений белков с углеводами. От внутренней, чисто белковой оболочки (капсида) она отделена тонким — всего в две молекулы толщиной — слоем липидов. Внутри капсида заключена вирусная РНК (не показана), единственная цепочка которой состоит примерно из 13 000 нуклеотидов. В столь ограниченном объеме уместилась бы информация максимум о 10–15 индивидуальных белках, но нужно ведь оставить место и на регуляторную часть (промотор), которая заставит клетку без устали считывать вирусные гены.


1–2. Вирусные белки
3. РНК
Проявляется в виде крапчатости и беловатых или желтоватых штрихов на листьях, окраска их становится светлой по сравнению с темнозелеными здоровыми листьями
Один из наиболее просто устроенных вирусов — вирус мозаики подорожника (RMV), поражающий многие виды растений, в том числе и культурных. Как и у многих других вирусов, его геном представлен единственной цепочкой РНК. Как известно, эта молекула имеет спиральную форму, и вдоль нее, тоже по спирали, располагаются молекулы вирусных белков. В результате образуется наборный чехлик-трубочка с молекулой РНК в качестве стержня. Сборка таких трубочек идет самопроизвольно, как кристаллизация, не требуя участия ферментов или иных активных факторов. Достаточно, чтобы в растворе были готовые вирусные белки и какая-нибудь однонитчатая молекула РНК — необязательно даже вирусная. При этом длина трубочек ограничена только длиной РНК и, в принципе, может быть любой (обычная вирусная частица RMV содержит 2100 молекул белка). Это свойство привлекло к вирусам мозаик внимание нанотехнологов, надеющихся таким образом собирать нанокабели.


1–3. Вирусные белки
4. Молекулы ICAM-1
Острая респираторная вирусная инфекция
Риновирусы впервые обнаружены в 1956 году. Они становятся виновниками не только острых респираторных вирусных инфекций (ОРВИ), но иногда и более серьезных заболеваний, включая астму, отиты, а также аллергические реакции у детей


1. Гликопротеидные шипы
2. Липидная оболочка
3. Белки капсида и матрикса
4. РНК
Синдром приобретенного иммунодефицита
Сто одежек, или Апофеоз упаковки
| ← Предыдущая глава | Глава 1.6 | Следующая глава → |
| Вирусы — неклеточные формы жизни | ||

Ви́рус (от лат. virus — яд) — микроскопическая частица, состоящая из белков и нуклеиновых кислот и способная инфицировать клетки живых организмов. Вирусы являются облигатными паразитами — они не способны размножаться вне клетки. В настоящее время известны вирусы, размножающиеся в клетках растений, животных, грибов и бактерий (последних обычно называют бактериофагами). Обнаружен также вирус, поражающий другие вирусы (Вирусы тоже болеют вирусными заболеваниями).
Вирусы представляют собой молекулы нуклеиновых кислот (ДНК или РНК), заключённые в защитную белковую оболочку (капсид). Наличие капсида отличает вирусы от других инфекционных агентов, вироидов. Вирусы содержат только один тип нуклеиновой кислоты: либо ДНК, либо РНК. Ранее к вирусам также ошибочно относили прионы, однако впоследствии оказалось, что эти возбудители представляют собой особые белки и не содержат нуклеиновых кислот.
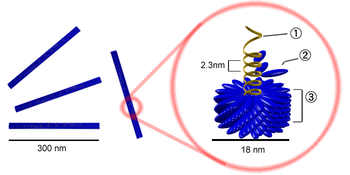

Роль вирусов в биосфере
Вирусы являются одной из самых распространённых форм существования органической материи на планете по численности: воды мирового океана содержат колоссальное количество бактериофагов (около 10 11 частиц на миллилитр воды), их общая численность в океане — около 4 х 10 30 , а численность вирусов (бактериофагов) в донных отложениях океана практически не зависит от глубины и всюду очень высока [1]. В океане обитают сотни тысяч видов (штаммов) вирусов, подавляющее большинство которых не описаны и тем более не изучены [2][3]. Вирусы играют важную роль в регуляции численности популяций животных.
Вирусные частицы (вирио́ны) представляют собой белковую капсулу — капсид, содержащую геном вируса, представленный одной или несколькими молекулами ДНК или РНК. Капсид построен из капсомеров — белковых комплексов, состоящих в свою очередь из протомеров. Нуклеиновая кислота в комплексе с белками обозначается термином нуклеокапсид. Некоторые вирусы имеют также внешнюю липидную оболочку. Размеры различных вирусов колеблются от 20 нм (пикорнавирусы) до 500 нм (мимивирусы). Вирионы часто имеют правильную геометрическую форму (икосаэдр, цилиндр). Такая структура капсида предусматривает идентичность связей между составляющими её белками, и, следовательно, может быть построена из стандартных белков одного или нескольких видов, что позволяет вирусу экономить место в геноме.
Фазы вирусной инфекции [ править ]
Условно процесс вирусного инфицирования в масштабах одной клетки можно разбить на несколько взаимоперекрывающихся этапов:
Классификация Балтимора и жизненные циклы вирусов [ править ]
Нобелевский лауреат, биолог Дэвид Балтимор, предложил свою схему классификации вирусов, основываясь на различиях в механизме продукции мРНК и связанных с этим особенностях жизненного цикла вирусов. .Эта система включает в себя семь основных групп:
- (VII) Вирусы, содержащие двуцепочечную ДНК и имеющие в своем жизненном цикле стадию синтеза ДНК на матрице РНК, ретроидные вирусы (например, вирус гепатита B).
История изучения вирусов [ править ]
В 1901 г. было обнаружено первое вирусное заболевание человека — жёлтая лихорадка. Это открытие было сделано американским военным хирургом У. Ридом и его коллегами.
В 1911 г. Фрэнсис Раус доказал вирусную природу рака — саркомы Рауса (лишь в 1966 г, спустя 55 лет, ему была вручена за это открытие Нобелевская премия по физиологии и медицине).
В последующие годы изучение вирусов сыграло важнейшую роль в развитии эпидемиологии, иммунологии, молекулярной генетики и других разделов биологии. Так, эксперимент Херши-Чейз стал решающих доказательством роли ДНК в передаче наследственных свойств. В разные годы еще как минимум шесть Нобелевских премий по физиологии и медицине и три Нобелевских премии по химии были вручены за исследования, непосредственно связанные с изучением вирусов.
В 2002 году, в университете Нью-Йорка был создан первый синтетический вирус (вирус полиомиелита).
Информация
Добавить в ЗАКЛАДКИ| Поделиться: |
КапсидВирусный капсид построен из полипептидных цепей (разных белков), сложенных в несколько слоев. У отдельных вирусов капсид окружен дополнительной мембраной, построенной из сложных вирусоспецифических белков (гликопротеидов) и двойного слоя липидов, идентичных липидам плазматической мембраны клетки-хозяина. Эту вирусную мембрану часто называют супер-капсидом.[ . ] Функциональное значение капсида определается тем, что он предохраняет вирусный геном (нуклеиновую кислоту) от повреждений, а также содержит рецепторы, обеспечивающие адсорбцию вирусных частиц на поражаемых ими клетках.[ . ] Белок ВТМ является единственным капсидиым белком вирусов растений, для которого полностью выяснена аминокислотная последовательность. Частичные последовательности установлены также для ВЖМТ Харрисом и- Хиндли [710]. Оказалось, что специфическое расщепление белка ВЖМТ трипсином приводит к образованию четырех довольно больших пептидов, которые плохо растворимы. Это и явилось препятствием на пути к определению полной аминокислотной последовательности белка ВЖМТ. Однако специфичность действия трипсина можно изменить. Методы гидролиза белков рассмотрены Хиллом [778].[ . ] ДНК-содержащие вирусы имеют либо собственные ферменты репликации (в капсиде), либо в их геноме закодирована информация о синтезе вирусных ферментов, обеспечивающих репликацию вирусной нуклеиновой кислоты. Количество этих ферментов различно в применении к разным вирусам. Например, в геноме бактериального вируса Т4 закодирована информация о синтезе около 30 вирусных ферментов. Далее геном крупных вирусов кодирует нуклеазы, разрушающие ДНК клетки-хозяина, ,а также белки, воздействие которых на клеточную РНК-полимеразу сопровождается тем, что «обработанная таким образом РНК-полимераза транскрибирует на разных стадиях вирусной инфекции разные вирусные гены. Напротив, малые по размерам ДНК-содержащие вирусы в большей мере зависят от ферментов клеток-хозяев. Например, синтез ДНК аденовирусов обеспечивается клеточными ферментами.[ . ] Известна полная последовательность 158 аминокислот в полипептидных цепях капсидиого белка ВТМ [23, 1797, 1928] и определена полная аминокислотная последовательность белков многих природных штаммов и искусственно полученных мутантов. Эти исследования внесли важный вклад в установление универсальной природы генетического кода и в наше понимание химических основ мутаций.[ . ] Частицы (или вирионы) большинства известных фагов имеют форму сперматозоида. Они состоят из головки (или капсида) и отростка. Наряду с этим есть фаги, которые состоят из одной головки, без отростка, и фаги, имеющие форму палочки (палочковидные или нитевидные фаги).[ . ] Как уже отмечено, репродукция вирусов происходит только в живых клетках, т. е. синтез как вирусного генома (нуклеиновой кислоты), так и белков капсида происходит только в клетках. В клетках осуществляется также и сборка из вирусной нуклеиновой кислоты и белков новых инфекционных вирусных частиц.[ . ] Вирусы — внутриклеточные паразиты. Могут проявлять свойства живых организмов, только попав внутрь клетки. Простые вирусы (например, вирус табачной мозаики) состоят из молекулы нуклеиновой кислоты и белковой оболочки — капсида. Вирус подавляет существующие в клетке процессы транскрипции и трансляции. Он использует их для синтеза собственных нуклеиновой кислоты и белка, из которых собираются новые вирусы.[ . ] Одиночные вирусы тех или иных видов представляют собой образования различной формы (округлой, палочковидной или другой формы), внутри которых содержится нуклеиновая кислота (ДНК или РНК), заключенная в белковую оболочку (капсид).[ . ] Что касается взаимодействия вирусных капсидных белков и нуклеиновых кислот при формировании вирусных частиц, то мысль о том, что сборка белковых оболочек мелких вирусов является спонтанным процессом, зависящим от свойств субъединиц капсидиого белка, была подтверждена экспериментами на палочкообразных вирусах [539] и мелких изометрических вирусах [76]. В этих экспериментах изолированные РНК и белковые субъединицы воссоединялись в соответствующих условиях in vitro с образованием интактных инфекционных вирусных частиц.[ . ] У некоторых РНК-содержащих вирусов в геноме закодированы РНК-зависимые транскриптазы (обратные транскриптазы). Поэтому репликация генома РНК-содержащих вирусов катализируется собственной обратной транскриптазой, которая упаковывается в капсиде при каждой репликации вирусов в клетке-хозяине. Такие РНК-содержащие вирусы получили название ретровирусов (от лат. retro — возврат назад).[ . ] Крик и Уотсон отметили, что если один и тот же тип взаимодействия между субъединицами повторяется многократно при построении вирусной частицы, то белковые молекулы должны быть правильно упакованы вокруг молекулы РНК. Существует ограниченное число способов упаковки субъединиц в вирусном капсиде. Большинство вирусов имеет либо палочкообразную, либо сферическую форму. Число белковых субъединиц, располагающихся в виде спирали в частицах палочкообразных вирусов, теоретически неограниченно.[ . ] Вирусная инфекция человека (животных) начинается с того, что инфицирующий вирус связывается с цитоплазматической мембраной клеток, после чего в результате эндоцитоза проникает внутрь клеток. Внутри клеток вирусные частицы оказываются или в ядре или в цитозоле, где их нуклеиновая кислота освобождается от капсида. Например, ДНК вируса герпеса человека освобождается от капсида в ядре. Там же происходит и сборка новых нуклеокапси-дов, после чего они покидают клетку. Напротив, РНК некоторых РНК-содержащих вирусов освобождается от капсида в цитозоле, после чего она транслируется на рибосомах клетки-хозяина с образованием кодируемой вирусом РНК-полимеразы. Последняя синтезирует РНК, отдельные из копий которой выполняют роль мРНК, транслируемой в мембранные н капсидные белки.[ . ] Следовательно, бактериальные вирусы по отношению к плазмидам являются прогрессивными структурами, которые в ходе эволюции приобрели специализированные функции, необходимые для осуществления сложного цикла — воспроизводства инфекционных вирусных частиц и для существования за пределами клетки-хозяина (во внеклеточной форме). Первый бактериальный вирус, вероятно, возник, когда у плазмид появился ген (гены), кодирующий белок (белки) капсида.[ . ] После того как было показано, что генетический код представляет собой последовательность триплетов оснований в нуклеиновой кислоте, каждый из которых определяет одну аминокислоту в белке, выяснилось следующее: большинство вирусов содержит значительно больше генетической информации, чем требуется для кодирования белка или белков, обнаруженных в составе вирусной частицы. Например, многие вирусы растений содержат молекулу РНК с молекулярной массой 2-10е дальтон. Этого достаточно, чтобы, кроме капсидиого белка, кодировать еще 5—8 белков средней молекулярной массы. Эти белки, по-видимому, нужны для размножения вируса и синтезируются в инфицированной клетке. По аналогии с результатами исследований на вирусах животных и вирусах бактерий можно предположить, что одним из таких белков является, вероятно, вирусоспецифичная РНК-синтетаза. Выделение и изучение свойств этих некапсидньтх белков является предметом дальнейших исследований.[ . ] В литературе отсутствуют данные о том, что РНК вирусов растений играет какую-либо роль в связывании белка антитела с вирусом. В условиях, при которых происходит ВТМ-инактивация целого вируса, аптисыворотка не оказывает выраженного влияния на инфекционную PIIK ВТМ [630]. В опытах с ■у-глобу-липовыми фракциями не отмечалось повреждения РНК, хотя инактивация целого вируса имела место. Эти результаты позволяют с очевидностью предположить, что белок антител инактивирует вирус непосредственно, а не косвенно путем какого-то воздействия на растение-хозяина. ВТМ, частично деградированный под действием лаурилсульфата натрия, чувствителен к рибоиуклеазе, поскольку часть РНК оказывается при этом обнаженной. Кроме того, такой препарат инактивируется антисывороткой, что, по-видимому, объясняется образованием комплекса между антителами и сохранившейся частью капсидиого белка [1397]. Связывание антител с вирусным белком может предотвратить освобождение РНК из белковой оболочки на ранних стадиях инфекции или препятствовать адсорбции вируса па определенных структурах внутри клетки.[ . ] Обычно принято считать, что все белковые субъединицы, из которых построена оболочка палочкообразных или мелких изометрических вирусов растений, идентичны. Однако эта однородность не продемонстрирована достаточно убедительно ни для одного вируса. Наиболее вероятно, что это утверждение справедливо для таких палочкообразных вирусов, как ВТМ. Очищенные иренараты вируса крапчатости бобов фасоли и вируса желтой мозаики коровьего гороха (ВЖМКГ) обладают электрофоретической гетерогенностью . Электрофорез в полиакриламидном геле в 8 М мочевине позволяет выявить в препарате белка, экстрагированного из иптактного ВЖМКГ 67%-ной уксусной кислотой, 6 основных и 2 минорные полосы [1534]. Природа этих фракций пока не установлена, по вполне возможно, что это одна и та нее химическая субъединица в различных физических состояниях, а не ряд разных полипептидов. Некоторые мелкие вирусы животных содерзкат более одного тина структурных белков. Вероятно, то же будет обнаружено и для некоторых вирусов растений. Так, исследования последнего времени позволяют предположить, что в состав капсида вируса мозаики коровьего гороха, вероятно, входят по крайней мере два различных белка (Щеферд, личное сообщение).[ . ] Обратимое окисление рибоиуклеазы перекисью водорода подавляет как ее ферментативную активность, так и способность ингибировать заражение ВТМ, но при этом фермент сохраняет способность образовывать комплекс ■с ВТМ [337J. Кастерман и Джинер [337] в качестве альтернативы инактивации в результате образования комплекса высказали мысль о том, что в процессе инфекции ВТМ проходит какую-то первоначальную стадию, когда вирус чувствителен к рибонуклеазе. Это подтверждается тем, что инфекционная РНК ВТМ очень чувствительна к инактивации ферментом. Концентрации фермента, в 1000 раз большие, чем те, при которых инактивируется РНК ВТМ, не оказывают влияния на иитактиый вирус. При инфильтрации разбавленных растворов рибоиуклеазы в листья табака [336, 337] было обнаружено, что это введение перед заражением блокировало развитие ВТМ. Инфильтрация после заражения проявлялась в постепенном ослаблении эффективности действия фермента в течение 2 ч; через 6 ч фермент не оказывал никакого влияния на развитие ВТМ. В листьях, которые просто погружали в раствор фермента на один час и затем отмывали перед заражением, размножение вируса протекало нормально. Вероятно, на недавно освобожденную из капсида инфекционную РНК можно воздействовать ферментом при ■его добавлении только на очень ранних стадиях инфекции.[ . ] Не приходится удивляться, что при таких вводных природа вирусов первое время была совершенно загадочна. Вирус мог оказаться мельчайшим живым организмом, а мог и просто ядовитой молекулой. Лет 100 назад биологи вполне допускали и то и другое. В 1935 году американский биохимик Уэнделл Стэнли ухитрился не только выделить из табачного сока вирус табачной мозаики, но и кристаллизовать его — так, как если бы это была обычная молекула. В то время это произвело сенсацию, тем более что под микроскопом вирусы на тот момент еще никто не видел. Неудивительно, что, получив этот результат, Стэнли посчитал вирус просто белковой молекулой, пусть и имеющей кое-какие особые свойства… …Постепенно биологам становилось ясно, что вирус…— это мелкий (не больше 200 нанометров) инфекционный агент, не обладающий автономностью и не способный размножаться делением, в отличие от живых клеток… Любой вирус обязательно содержит помимо белка нуклеиновую кислоту, но только одного типа — или ДНК, или РНК. Последнее резко отличает вирусы от живых клеток, в состав которых… всегда входят оба типа нуклеиновых кислот… В любом случае такие взгляды подразумевали, что вирусы — это по сути своей нечто более простое, чем клетки. Гораздо, качественно более простое. Иная ступень устройства природы, если угодно. Все гигантские вирусы оказались ДНК-содержащими, причем ДНК у них двуцепочечная. Прочтение генома мимивируса показало, что этот геном состоит из 1,18 миллиона пар нуклеотидов и включает 979 генов, кодирующих белки… Он приближается по величине к геномам клеточных организмов — и не просто приближается, а вполне достигает их размеров… Но ведь и мимивирус не уникален. Исследования быстро показали, что гигантских ДНК-содержащих вирусов в природе не так уж мало. За прошедшие годы было открыто еще несколько близких друг к другу родов гигантских ДНК-содержащих вирусов. Они получили разнообразные названия: марсельвирусы, мамавирусы, мегавирусы, пандоравирусы, молливирусы, питовирусы и даже моумоувирусы… Впрочем, полного аппарата трансляции, способного к самостоятельной работе, нет и у клоснойвируса. У него полностью отсутствуют гены, кодирующие рибосомную РНК и рибосомные белки… — это, как мы уже говорили, одно из самых главных различий между вирусами и клетками. По этому признаку никаких переходных форм между ними пока не видно. Так или иначе, открытие гигантских вирусов смело можно назвать одним из важнейших в современной биологии. Оно целиком датируется XXI веком. Открытие гигантских вирусов заметно изменило наши представления о структуре живой природы в целом. Конечно, никто не утверждает, что пандоравирус устроен сложнее кита-полосатика. Верхний предел сложности (даже чисто генетической) у клеточных организмов явно гораздо выше; это видно хотя бы по размерам их геномов. Но вот сказать, что любая клетка устроена сложнее любого вируса, теперь нельзя никак.
…Бросается в глаза, что есть как минимум две важные группы генов, свойственных клеткам, но не свойственных вирусам. Это гены энергетического обмена и гены, ответственные за создание рибосом. У подавляющего большинства вирусов никаких генов энергетического обмена нет вообще, а если даже они есть, то кодируют только отдельные ферменты, но не полную систему синтеза АТФ … Судя по биоинформатическим данным, последний общий предок всех клеточных организмов имел как минимум 34 рибосомных белка, и эти белки (вместе с кодирующими их генами) сохранились у всех бактерий, архей и эукариот. У вирусов же нет ни одного из них.
Простые (безоболочечные) вирусы состоят из нуклеиновой кислоты и белка и представляют собой нуклеопротеиды или нуклеокапсиды. Сложные (оболочечные) вирусы кроме нуклеиновой кислоты и белка содержат также липиды и углеводы. Нуклеиновые кислоты. Клетки всех живых организмов содержат два типа нуклеиновых кислот — ДНК и РНК. Их структура относительно однообразна: ДНК представляет собой двуспиральную молекулу, РНК — односпиральную молекулу. Функция ДНК заключается в хранении и репликации наследственной информации, таким образом, она является клеточным геномом. РНК представлена в клетке в трех формах: информационной (иРНК), рибосомальной (рРНК) и транспортной (тРНК). Каждая из них выполняет определенную функцию: иPHК образуется в результате транскрипции генома и передает информацию с генома на белоксинтезирующий аппарат клетки, рРНК является структурным компонентом рибосом, тРНК доставляет аминокислоты белоксинтезирующему аппарату клетки. В отличие от клетки вирусы содержат один тип нуклеиновой кислоты — или ДНК, или РНК. Каждая из них выполняет функцию вирусного генома. Структура нуклеиновых кислот у разных вирусов весьма разнообразная. По количеству цепей они бывают одно — и двуспиральными, по форме — линейными и кольцевыми (циркулярными), а также непрерывными и фрагментированными. Содержание нуклеиновой кислоты в вирионе различных вирусов составляет от 1 % у ортомиксо- и парамиксовирусов до 32 % у парвовирусов и не коррелирует ни с систематическим положением вируса, ни со степенью сложности его организации. Вирусные ДНК. Молекулярная масса ДНК различных вирусов варьирует в широких пределах: от 2 МД у цирко — и парвовирусов до 375 МД у поксвирусов. Самые большие геномы содержат до нескольких сотен генов, самые маленькие — несколько генов. По структуре молекулы ДНК бывают одно — и двуспиральными, линейными и кольцевыми. У вирусов с двуспиральными ДНК информация обычно закодирована на обеих спиралях, что говорит о максимальной экономии генетического материала. Большинство нуклеотидных последовательностей в молекуле ДНК встречается однократно. Однако в концевом фрагменте линейных ДНК возможно наличие ее начального участка в виде повтора, который бывает прямым или инвертируемым. Благодаря таким повторам молекулы ДНК могут приобретать циркулярную форму, которая обеспечивает их устойчивость к эндонуклеазам. Кроме того, стадия образования циркулярной формы обязательна для интеграции вирусной ДНК с геномом клетки. Вирусные РНК. Молекулярная масса вирусных РНК варьирует в пределах от 4—5 МД у нодавирусов до 32 МД у реовирусов. По предложению Балтимора (1971) вирусы с односпиральными РНК из-за различий в функциях генома было принято разделять на две подгруппы. У вирусов первой подгруппы вирусный геном обладает функциями иРНК и их условно обозначают как плюс-нитевые вирусы, или вирусы с позитивным геномом. У вирусов второй группы РНК не обладает функцией иРНК. На ней, как на матрице, синтезируется комплементарная молекула. Это происходит только в присутствии вирусного белка — фермента транскриптаза, который обязательно находится в структуре минус-нитевых вирусов (в клетках — ее аналога нет). Существуют вирусы, которые содержат как плюс-нитевые, так и минус-нитеные РНК-гены, так называемые амбигюлирные РНК. Белки. Белки всех известных в настоящее время вирусов позвоночных являются основными компонентами вирионов и составляют от 57 до 90 % массы вириона. По аминокислотному составу вирусные белки принципиально не отличаются от состава белков животных. В геноме вирусов кодируются две группы белков: структурные, которые входят в состав вирионов потомства, и неструктурные, участвующие в репродукции вируса на разных этапах, но не входящие в состав вирионов. Структурные белки в составе вириона варьируют в широких пределах, что зависит от сложности организации вириона. Простые вирусы животных содержат 3—4 белка, сложные, например вирусы оспы, — более 30. Среди структурных белков различают две группы — капсидные и пепломеры. Первая группа включает белки, которые формируют капсид (от греч. capsa — вместилище), окружающий нуклеиновую кислоту, а также геномные белки и ферменты; вторая группа — белки суперкапсидной оболочки, называемой пеплос (от греч. peplos — покров, мантия). Простые вирусы содержат только капсидные белки, сложные — и капсидные, и пепломеры. Белки в составе вирусного калсида называются капсомерами. Основной их функцией является защита вирусного генома от неблагоприятных воздействий внешней среды. Они представляют собой идентичные полипептидные цепи (белковые субъединицы), которые обладают способностью к самосборке. Сборка капсида из субъединиц запрограммирована в первичной структуре белка и происходит самопроизвольно или при взаимодействии с нуклеиновой кислотой. Принцип субъединичности в строении капсида — уникальное свойство капсидных белков, благодаря которому достигается огромная экономия генетического материала. Кроме того, в механизме самосборки заложена возможность контроля за белковыми субъединицами: дефектные и, чужеродные белковые цепи не включаются в капсид. Принцип самосборки характерен только для простых вирусов. Сложные вирусы сборку осуществляют по более сложному многоступенчатому механизму. Однако отдельные ее этапы (формирование капсидов и нуклеокапсидов) основаны на самосборке. Суперкапсидные белки (пепломеры) находятся в липопротеидной оболочке сложных вирусов. Они либо пронизывают липидный бислой вириона, либо не доходят до его внутренней поверхности. Являясь типичными внутримембранными белками, они, как правило, гликозилированы (гликопротеиды), т. е. к молекуле белка в определенных местах прикреплены углеводные цепи. Гликозилирование осуществляют клеточные ферменты, поэтому один и тот же вирус, но реплицирующийся в разных клетках, может иметь разные углеводные остатки по составу углеводов, длине углеводной цепи, месту прикрепления к белку. Неструктурные белки менее изучены. Существуют определенные трудности в их выделении из зараженных клеток и очистки от клеточных белков. К ним относятся: предшественники вирусных белков, которые существуют в зараженной клетке очень непродолжительное время, а затем нарезаются; ферменты синтеза РНК. и ДНК — полимеразы; регуляторы стадий репродукции вирусов; ферменты, модифицирующие вирусные белки — протеиназы и протеинкиназы. Липиды и углеводы. В состав вирионов всех сложных (оболочечных) вирусов позвоночных кроме нуклеиновой кислоты и белков входят липиды и углеводы. Состав липидов вирионов сходен с липидным составом клетки хозяина: примерно 50—60 % составляют фосфолипиды и 20—30 % — холестерин. У отдельных представителей липидов содержится до 20—35 % от массы вириона (ортомиксо-, ретро-, буньявирусы). Липиды обнаружены только в суперкапсидной оболочке вирионов и имеют клеточное происхождение. Это связано с тем, что оболочечные вирусы формируются путем почкования на плазматической мембране клеток. Поэтому суперкапсидная оболочка вирионов представляет собой мембрану клетки-хозяина, модифицированную за счет встроенных в нее вирусных белков — пепломеров. Липидный компонент стабилизирует структуру вирусных частиц, поэтому их обработка детергентами или липазами приводит к потере инфекционности. Исключение составляют вирусы оспы. У них липиды не образуют дифференцированной оболочки. Обработка вирусов осповакцины жирорастворителями не приводит к потере инфекционной активности или каким-либо другим структурным изменениям вириона. Углеводы находятся в вирионах в виде гликопротеинов, встроенных в суперкапсидный слой, а также гликолипидов и имеют клеточное происхождение. У отдельных представителей вирусов позвоночных содержание углеводов доходит до 7—8 % от массы вириона (ортомиксо-, тогавирусы). Химический состав их полностью определяется клеточными ферментами, которые обеспечивают перенос и присоединение сахарных остатков. В вирионах в основном обнаруживают фруктозу, сахарозу, маннозу, галактозу, нейраминовую кислоту, глюкозамин. Углеводы являются каркасом для локальных участков гликопротеидов, обеспечивают сохранение конформации белковых молекул и защищают от действия протеаз. Компоненты клетки-хозяина. В составе вирионов могут обнаруживаться некоторые компоненты клетки-хозяина. Это могут быть белки или цельные клеточные структуры. Например, в составе некоторых оболочечных вирусов находится белок цитоскелета актин, в составе папиллома — и полиомавирусов содержатся клеточные гистоны, у ареновирусов обнаружены рибосомы. Клеточные компоненты могут включаться в вирион случайно или закономерно. Во втором случае они могут играть существенную роль в репродукции вирусов, как, например, гистоны для папиллома — и полиомавирусов. Структурная организация вирионов. Вирионы (вирусные частицы) по архитектуре подразделяются на два типа: имеющие внешнюю липопротеидную оболочку (сложные, или оболочечные, вирусы) и не имеющие такой оболочки (простые, или безоболочечные, вирусы). Капсомеры соединяются друг с другом строго определенным образом и образуют только два типа капсидов: спиральный и изометрический (квазиметрически й). Только такие способы укладки капсомеров вокруг нуклеиновой кислоты (типы симметрии) обеспечивают формирование энергетически экономичных структур с минимумом свободной энергии. Изометрические капсиды представляют структуры, в которых капсомеры соединяются между собой в правильные многогранники, в центре которых расположен геном. Такой способ укладки капсомеров называется кубическим типом симметрии, а многогранник — икосаэдром. Это означает, что он симметричен в трех взаимно перпендикулярных направлениях и его линейные размеры вдоль прямоугольных осей идентичны. Обычно изометрические капсиды состоят из 60 (или кратных 60) геометрически идентичных элементов, которые имеют 12 вершин, 20 граней и 20 ребер. Так, капсид вируса полиомиелита образован 60 белковыми структурными единицами, каждая из которых состоит из четырех полипептидных цепей (химических единиц) — vp1, vp2, vp3, vp4 (virion protein). Наиболее сложно структурно организованы вирусы оспы. Их сердцевина (нуклеоид), содержащая вирусную ДНК в составе нуклеотида, имеет форму двояковогнутого кольца и окружена двумя линзообразными латеральными тельцами. Капсид окружен дополнительными внутренними белковыми структурами — вирусным матриксом. По архитектуре вирусы оспы отличаются от всех других вирусов позвоночных и их принято называть вирусами со сложным типом симметрии. Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter. Читайте также:
 Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу. Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу.
Copyright © Иммунитет и инфекции
|