Транскрипция и трансляция у вирусов
После расшифровки генетического кода встал вопрос: каким образом осуществляется перенос информации с ДНК на белок? Биохимическими исследованиями было установлено, что основная масса ДНК в клетке локализована в ядре, тогда как синтез белка идет в цитоплазме. Это территориальное разобщение ДНК и синтеза белка обусловило поиски посредника. Поскольку синтез белка шел с участием рибосом, то на роль посредника была выдвинута РНК. Была создана схема, иллюстрирующая направление потока генетической информации в клетке:
ДНК → РНК → белок
Она получила название центральной догмы молекулярной биологии. Ф. Крик постулировал, что синтез макромолекул по этой схеме осуществляется по матричному принципу. На доказательство правильности этого постулата потребовались многие годы.
Схема процесса транскрипции

Вначале предполагалось, что роль посредника выполняет рибосомальная РНК (“один ген — одна рибосома — один белок”). Однако в скором времени выяснилась несостоятельность такого предположения. Было показано, что в процессе белкового синтеза количество рибосом не изменяется, т.е. новая РНК не синтезируется и, следовательно, новая информация не поступает. Вскоре в составе рибосом была обнаружена фракция нестабильной РНК, молекулы которой непрочно удерживаются на рибосоме с помощью катионов Mg. Методом молекулярной гибридизации было показано, что молекулы этой РНК являются копиями определенных участков ДНК. Она получила название матричной, или информационной РНК. Ее также называли раньше РНК-посредник и мессенджер-РНК. Комплементарность этих молекул определенным участкам ДНК говорила о том, что они синтезируются по матричному типу на ДНК.
Постепенно был выяснен весь путь переноса информации от ДНК к белку. Он состоит из двух этапов: транскрипции и трансляции. На этапе транскрипции происходит считывание и перенос генетической информации с ДНК на иРНК. Процесс транскрипции протекает в три стадии: инициации, элонгации и терминации. Информация считывается только с одной цепи ДНК (+ цепь), так как исходя из свойств генетического кода, комплементарные участки ДНК не могут кодировать структуру одного и того же белка из-за отсутствия комплементарной вырожденности кода. Ведет транскрипцию фермент РНК-полимераза, состоящий из четырех субъединиц (ααββ') и не обладающий специфичностью в отношении источника ДНК. На начальном этапе транскрипции — инициации — к ферменту присоединяется пятая субъединица, так называемый s-фактор, который осуществляет узнавание специфического участка ДНК, промотора. Промоторы не транскрибируются. Узнаются они s-фактором по наличию в них специфической последовательности нуклеотидов. В бактериальных промоторах она называется блоком Прибнова и имеет вид ТАТААТ (с небольшими вариациями). К промотору присоединяется фермент РНК-полимераза. Рост цепи иРНК идет в одном направлении, скорость транскрипции равняется ≈ 45-50 нуклеотидов в 1 секунду. На этапе инициации синтезируется только короткая цепочка из 8 нуклеотидов, после чего s-фактор отделяется от РНК-полимеразы и начинается этап элонгации. Наращивание цепи иРНК ведет уже белок-тетрамер. Участок, с которого считывается информация, называется транскриптоном. Он заканчивается терминатором — специфической нуклеотидной последовательностью, играющей роль stop-сигнала. Дойдя до терминатора, фермент РНК-полимераза прекращает работу и с помощью белковых факторов терминации отделяется от матрицы.

Структура молекулы тРНК
В бактериальных клетках образующиеся молекулы иРНК могут сразу выполнять роль матриц для синтеза белка, т.е. транслироваться. Они соединяются с рибосомами, к которым одновременно молекулы транспортных РНК (тРНК) доставляют аминокислоты. Цепочки транспортных РНК состоят примерно из 70 нуклеотидов. Однонитиевая молекула тРНК имеет участки комплементарного спаривания, в составе которых находятся активные центры: участок узнавания тРНК ферментом тРНК-синтетазой, присоединяющим к тРНК соответствующую активированную аминокислоту; акцептор — участок, к которому присоединяется аминокислота, и антикодоновая петля.
Схема синтеза белка (трансляция)

Антикодон — это триплет, комплементарный соответствующему кодону в молекуле иРНК. Взаимодействие кодон-антикодон идет по типу комплементарного спаривания, во время которого происходит присоединение аминокислоты к растущей белковой цепи. Инициирующим кодоном в составе разных иРНК является кодон AUG, соответствующий аминокислоте метионину. Поэтому первой к матрице подходит тРНК с антикодоном UAC, соединенная с активированной аминокислотой метионином. Ферменты, активирующие аминокислоты и соединяющие их с тРНК, называются аминоацил-тРНК-синтетазы. Все этапы биосинтеза белка (инициация, элонгация, терминация) обслуживаются белковыми факторами трансляции. У прокариот их по три на каждый этап. В конце матрицы иРНК находятся нонсенс-кодоны, которые не считываются и знаменуют собой конец трансляции.
В геноме многих организмов, от бактерий до человека, обнаружены гены и соответствующие им тРНК, осуществляющие нестандартное считывание кодонов. Это явление получило название неоднозначности трансляции.

Оно позволяет избежать негативных последствий ошибок, возникающих в структуре молекул иРНК при транскрипции. Так, при появлении внутри молекулы иРНК нонсенс-кодонов, способных преждевременно прекратить процесс транскрипции, включается механизм супрессии. Он состоит в том, что в клетке появляется необычная форма тРНК с антикодоном, комплементарным нонсенс-кодону, чего в норме быть не должно. Ее появление является результатом действия гена, осуществляющего замену основания в антикодоне тРНК, близким по составу к нонсенс-кодону. В результате такой замены нонсенс-кодон считывается как обычный значащий кодон. Подобные мутации получили название супрессорных, т.к. они подавляют изначальную мутацию, которая привела к появлению нонсенс-кодона.
Читайте также другие статьи темы 6 "Молекулярные основы наследственности":
Перейти к чтению других тем книги "Генетика и селекция. Теория. Задания. Ответы":
10 О бщая м едицинская вирусология
Таким способом проникают в клетку простые вирусы и многие слож ные вирусы .
Второй способ проникновения заключается в слиянии суперкапсидной оболочки вируса с клеточной мембраной, осуществляемой белками слияния, в результате чего внут ренняя часть вируса (его нуклеокапсид, сердцевина) оказы вается в цитоплазме клетки. Такой способ проникновения характерен для слож ных вирусов, обладающих F-белка- ми-слияния или другими гликопротеинами, выполняю щими их функции (например, гемагглютинин у вируса гриппа).
освобож дения его нуклеиновой кислоты такж е отличает ся у вирусов, проникающ их в клетку разными способами.
При способе проникновения вирусов путем слияния мембран депротеинизация вирионов начинается уж е при их проникновении с помощ ью ферментов клеточной мем браны и продолжается внутри клетки, как указано выше.
В результате депротеинизации происходит дезинтегра ция вириона. Освобожденная геномная нуклеиновая кис лота приобретает способность индуцировать репродукцию вирусов. У некоторы х вирусов освобож дение нуклеино вой кислоты бывает полным, но обычно в ассоциации с нуклеиновой кислотой остаются геномные (внутренние) белки, например, полимеразы и иногда — капсидные бел ки. В дальнейшем эти белки участвуют в процессе репро дукции и защите нуклеиновой кислоты от действия кле точных нуклеаз.
1 ЛАВА 1 . 1*С£Д£лИл U
4. Экспрессия ви русн ого геном а становится возм ож ной после высвобож дения вирусной нуклеиновой кисло ты, иногда требуется ее транспортировка в ядро клетки и взаимодействие с клеточным геномом.
Реализация генетической программы вируса начина ется с процесса транскрипции с последующ ей трансляци ей и репликацией вирусного генома, в результате чего образуются компоненты вируса — копии вирусного гено ма и структурные белки вируса.
Транскрипция — образование на матрице геномной нук леиновой кислоты комплементарных информационных РНК (и-РН К), необходимы х для последующ ей трансля ции и синтеза вирусных белков на рибосомах клетки.
Трансляция — процесс перевода генетической инфор мации, заложенной в и-РН К, на специфическую последо вательность аминокислот. Трансляция осущ ествляется и- РНК на клеточных рибосомах, на которы х синтез клеточ ных белков подавляется и транслируются вирусные белки.
Существует два способа формирования вирусных бел ков в зависимости от длины и-РНК. Короткие, моноцистронные и Р Н К кодирую т отдельный, обычно зрелый, ви русный белок. Длинные полицист ронные и-РНК, кото рые могут содержать всю информацию вирусного генома или ее часть, поступают на полирибосомы , на которы х транслируется один гигантский полипротеин-предшествен ник. Этот полипептид нарезается вирусными и клеточны ми протеазами на отдельные зрелые вирусные белки как неструктурные (обслуж иваю щ ие процесс репродукции ферменты и регуляторные белки), так и структурные. У не которых вирусов эти два способа формирования белков могут сочетаться.
Транскрипция и трансляция имеют свои особенности в зависимости от типа и строения вирусных нуклеиновых кислот.
Геномная ДНК > транскрипция > и-РНК > трансля ция > белок.
О бщ ая м едицинская вирусология
Если транскрипция происходит в ядре, как у больш ин ства ДНК-вирусов, то она осущ ествляется клеточной ДНКзависимой РН К -полимеразой (транскриптазой), если в цитоплазме, то транскрипцию выполняет вирусная транскриптаза, входящ ая в состав вириона. У ДНК -вирусов последовательно считы ваю тся участки генома, кодирую щие отдельные белки, образую тся короткие и-РН К, сна чала ранние и-РНК, затем поздние и-РНК. Соответствен но происходит трансляция на рибосомах сначала ранних неструктурны х, а затем поздних — структурных белков вируса.
2. У плюс-РН К вирусов геномная РНК одновременно является информационной РНК, поэтому стадия
транскрипции отсутствует и схема укорочена:
Геномная плюс-РНК > трансляция > белок.
П лю с-РН К, выполняющ ая функцию и-РН К, поступает на полирибосомы и полностью транслируется с образова нием гигантского полипептида-предшественника, который нарезается протеазами на отдельные белки — неструктур ные и структурные.
3. У м инус-РН К вирусов (с однонитевой и двунитевой РНК) синтез белка происходит по следующ ей схеме:
Геномная минус-РНК > транскрипция > и-РНК > транс ляция > белок.
Транскрипция осущ ествляется собственны ми транскриптазами вирусов, при этом возмож но образование как коротких, так и длинных и-РНК с последующ ей трансля цией индивидуальных зрелых белков или полипептидапредшественника.
Сущ ествуют и другие типы транскрипции и трансля ции, например у ретровирусов.
4. Рет ровирусы (возбудитель ВИЧ-инфекции, онкогенные вирусы) имеют диплоидный геном, состоя щий из двух идентичных молекул однонитевых плюс-РНК, и фермент РН К -зависимую ДНК-поли- меразу, называемую такж е обратной транскрипта зой или ревертазой.
Г лава 1. О с новные с ведения о вирусах
Ретровирусам свойственен уникальный, очень редко встречающийся у вирусов путь передачи генетической информации с РНК на ДНК. Подобный путь обнаружен также у вируса гепатита В и, предполож ительно, у виру са клещевого энцефалита.
Схема синтеза белка у ретровирусов следующ ая:
Геномная РНК > комплементарная ДНК (провирус) > транскрипция> и-РНК > трансляция > белок.
На матрице геномной плюс-РНК с помощ ью обратной транскриптазы синтезируется комплементарная ДНК (ми нус-нить). Затем вирусная РНК разрушается нуклеопротеазой и вместо нее достраивается вторая нить ДНК (плюснить) с участием клеточной ДНК -полимеразы . Образует ся двунитевая ДНК (несущ аяя генетическую информацию ретровируса), которая, приняв кольцевидную форму, ин тегрирует в хром осом у клетки в виде провируса. С этой провирусной ДНК происходит транскрипция и-РН К и далее трансляция белков.
Репликация вирусного генома заключается в синтезе на матрице исходной геномной нуклеиновой кислоты вируса множества копий — будущ их вирусных геномов. У боль шинства вирусов репликация происходит в ядре клетки, у некоторых — в цитоплазме. .Процесс начинается после накопления неструктурны х ранних белков, необходимых для обслуживания репликации.
Репликация вирусного генома имеет сходство с транс крибированием и-РН К, осущ ествляется вирусными или клеточными полимеразами (возмож но несколько модифи цированными). Главное отличие состоит в том, что при репликации вирусный геном всегда считывается полно стью и образующ иеся копии идентичны матричному ге ному. Как нам известно, при транскрипции и-РНК часто считываются отдельные гены или небольшие участки ви русного генома. Существует регуляция переключения про цессов репликации на транскрипцию и обратно, однако механизм такого переключения не вполне ясен.
Репликация имеет отличия у вирусов с различным ти пом генома.
О бщ ая медицинская вирусология
1. Репликация двунитевых ДНК-геномов осущ еств ляется с помощ ью клеточной ДНК-зависимой ДНКполимеразы (репликазы) по полуконсервативному типу, подобно клеточным ДНК.
2. Однонитевые плюс-РНК геномы реплицируются с помощ ью вирусиндуцированной РНК-полимера- зы. На исходной нити плюс-РН К синтезируется комплементарная минус-нить, образуется промежу точный репликативный комплекс, состоящ ий из двух нитей. На отделяющ ихся минус-РН К форми руются нити плюс-РНК, идентичные исходной ге номной плюс-РН К, происходит накопление многих копий генома.
3. Однонитевые минус-РНК геномы реплицируют ся с помощ ью входящ ей в состав вириона РНКзависимой РНК-полимеразы по тому же принци пу, что и плюс-РН К вирусы, т.е. через промеж у точный репликативный комплекс. В результате образуется множ ество копий минус-РН К геномов, идентичных исходному.
4. У ретровирусов для осущ ествления репликации геномной РНК необходимо прохождение тех же ста дий, что при транскрипции с обязательной интег рацией ДНК -провируса в хром осом у клетки. На матрице провирусной ДНК с помощ ью клеточной ДНК -зависимой РНК-полимеразы реплицируются копии однонитевых плюс-РНК — будущ их гено мов дочерней популяции ретровирусов.
Для ретровирусов характерно сочетание интегративной и продуктивной инфекции клеток. Если преобладает ин тегративная инфекция, провирус длительно сохраняется в зараженной клетке и при ее делении в составе хром осо мы переходит в дочерние клетки (наблюдается персистенция вируса).
Таким образом, в результате экспрессиии вирусного генома в инфицированной клетке накапливаются компо ненты вируса — копии генома и структурные белки. Син тез нуклеиновых кислот и белков вируса обычно проис
Г лава 1. О сновные с ведени я о вирусах
ходит в разных частях клетки и не одновременно, поэто му такой способ репродукции вирусов называет ся дизъ юнктивным или разобщ енным .
5. Ф орм ирование вирионов из отдельных компонен тов вируса у больш инства вирусов осущ ествляется в цитоплазме.
Простые вирусы образуются путем самосборки при вза имодействии нуклеиновой кислоты вируса и капсидных белков, при этом капсид формируется по спиральному или кубическому типу симметрии. Получаемая структура ста
бильна и называется нуклеокапсидом.
Сложные вирусы ф орм ирую тся в несколько этапов. Сначала образуются нуклеокапсиды, которы е взаимодей ствуют с модифицированными мембранами клетки (на ружными или внутренними, в том числе ядерной), одева ются суперкапсидной оболочкой, у некоторых вирусов под суперкапсидом формируется матриксный слой (М -белок).
6. В ы ход вирионов из клет ки происходит или при разрушении, лизисе клетки, или путем почкования.
Второй способ наблюдается у слож ны х вирусов, кото рые, почкуясь через мембраны клетки, одновременно при обретают суперкапсидную оболочку клеточного происхож дения со встроенными вирусными белками — гликопро теинами. При этом клетка, как правило, погибает не сразу, продолжая выделять новые поколения вирионов вплоть до истощения ее ресурсов.
1.2. Таксономия и классификация вирусов
Современная классиф икация вирусов является уни версальной и включает вирусы , поражающ ие человека и позвоночных ж ивотных, беспозвоночных, простейш их, ра стения, грибы, бактерии. Как уже указывалось, вирусы выделены в отдельное царство — Vira.






Общие принципы вирусологии
Цель дисциплины – познакомить студентов с вирусами и субвирусными агентами.
Задача дисциплины - сформировать у студентов представление о:
· месте вирусов и субвирусных агентов в природе
· строении вирусных частиц,
· разнообразии вирусных геномов,
· особенностях взаимодействия вирусов с клеткой и механизмами их репликации,
· природе субвирусных агентов – вироидов, сателлитных РНК и прионах,
IV. Место дисциплины в структуре ООП
Информация об образовательном стандарте и учебном плане:
· тип образовательного стандарта и вид учебного плана – специалитет
· направление подготовки – фармация и лечебное дело
Информация о месте дисциплины в образовательном стандарте и учебном плане:
Для освоения дисциплины необходимо освоение следующих дисциплин: цитологии, органической химии, биохимии, молекулярной биологии, иммунологии
Общая трудоемкость курса – 36 ак. часа
Форма промежуточной аттестации – зачет
V. Форма проведения дисциплины –
Форма проведения дисциплины – лекции
Форма текущего контроля – коллоквиум.
VI. Распределение трудоемкости по разделам и темам, а также формам проведения занятий с указанием форм текущего контроля и промежуточной аттестации:
Трудоемкость (в ак. часах) по формам занятий (для дисциплин) и видам работ (для практик)
Аудиторная работа (с разбивкой по формам и видам)
Наименование разделов и тем дисциплины/наименование разделов (этапов практики)
Раздел I. Общие сведения о вирусах
Тема 1. Краткие сведения о вирусах.
Тема 2. Вирусная инфекция
Тема 3. Структура генетического материала вирусов.
Тема 4. Структура вирусных частиц.
Раздел II. Взаимодействие вируса и клетки.
Тема 1. Проникновение вируса в клетку
Тема 2. РНК-содержащие вирусы.
Тема 3. ДНК-содержащие вирусы.
Коллоквиум по разделам 1 и 2
Раздел III. Синтез компонентов вируса.
Тема 2. Обратная транскрипция.
Тема 3. Синтез вирусных белков
Тема 4. Формирование вирусных частиц
Раздел IV. Онкогенные вирусы и медленные инфекции
Тема 1. РНК-содержащие онкогенные вирусы.
Тема 2. ДНК-содержащие онкогенные вирусы.
Тема 3. Медленные вирусные инфекции.
Тема 4. Прионные болезни
Коллоквиум по разделам 3 и 4
VII. Содержание дисциплины
Раздел I. Общие сведения о вирусах
Тема 1. Краткие сведения о вирусах.
Тема 2. Вирусная инфекция.
Стадии инфекционного цикла. Терминология. История изучения инфекционного цикла. Культивирование клеточных линий. Цитопатический эффект. Синцитиеобразование. Оценка инфекционности вирусов. Методы определения инфекционности. Метод бляшек. Метод конечного разведения. Отношение количества вирусных частиц к количеству бляшкообразующих единиц.
Оценка количества вирусных частиц: гемагглютинация, электронная микроскопия, определение активности вирусных ферментов, иммуноблоттинг, ИФА. Современные методы: использование флуоресцентных белков, ПЦР, секвенирование. Работы Эллис и Дельбрюк. Одиночный цикл размножения фага. Множественность инфекции.
Тема 3. Структура генетического материала вирусов
Классификация вирусов по Балтимору (в зависимости от типа генетического материала). ДНК-содержащие вирусы. Различные типы ДНК-геномов: непрерывный дцДНК, сегментированный дцДНК, оцДНК.
РНК-содержащие вирусы. Различные типы РНК-геномов: дцРНК, оцРНК (+), оцРНК (-). Реассортация сегментированных геномов. Двусмыленная РНК.
Генетика вирусов. Трансформация, трансфекция, мутагенез. Эксперимент Херши-Чейз. Генная терапия.
Тема 4. Структура вирусных частиц.
Общие принципы структуры вирусов. Терминология. Функции вирусных белков. Молекулярная организация вирионов.
Методы изучения структуры вирусных частиц: электронная микроскопия, рентгеноструктурный анализ, криоэлектронная микроскопия, ЯМР-спектроскопия.
Общие принципы структуры вирусов, симметрия. Спиральные вирионы (принципы спиральной симметрии, вирус табачной мозаики). Принципы симметрии сферических вирусов. Икосаэдрические вирусы. Строение оболочечных вирусов. Функции гликопротеинов. Сложные вирусы (аденовирусы, реовирусы, герпесвирусы). Бактриофаги.
Раздел II. Взаимодействие вируса и клетки.
Тема 1. Проникновение вируса в клетку.
Этапы взаимодействия вирусной частицы и клетки. Клеточные рецепторы, необходимые для вирусных частиц. Механизмы проникновения вируса в клетку. Влияние рН на процесс проникновения на примере вируса гриппа. Механизмы регуляции. Молекулярные механизмы проникновения филовирусов в клетку. Взаимодействие простых вирусов с клеточной мембраной. Роль корецепторов в инициации вирусной инфекции. Механизмы проникновения в клеточное ядро.
Тема 2. РНК-содержащие вирусы.
История изучения РНК. Терминология. Типы РНК-геномов. Общие принципы синтеза РНК в клетке. Особенности транскрипции и трансляции (+) РНК-содержащих вирусов. Созревание и выход вирионов из клетки. Репликация и транскрипция (-) РНК-содержащих вирусов с непрерывным геномом. Репликация и транскрипция (-) РНК-содержащие вирусы с сегментированным геномом. РНК-содержащие вирусы с двуцепочечной РНК. Синтез и трансляция вирусных мРНК. Репликация двуцепочечных РНК и созревание вирионов. Синтез РНК как источник генетической изменчивости.
Тема 3. ДНК-содержащие вирусы.
Общие принципы синтеза ДНК в клетке. Особенности репликации вирусных ДНК-геномов. Роль клеточных и вирус-специфических белков в репликации ДНК. Два механизма репликации дцДНК. Обезьяний вирус-40 (SV-40): механизм репликации, роль клеточных и вирусных белков. Парвовирусы: самозатравочный механизм инициации синтеза ДНК. Репликативная форма, надрез (nick) вирусной эндонуклеазой. Инверсия концевых повторов.
Аденовирусы: роль белков в инициации синтеза ДНК. Точка начала репликации (ori) вирусов.
Крупные ДНК-содержание вирусы: механизм репликации вируса простого герпеса и вируса оспы.
Регуляция синтеза вирусных ДНК
Раздел III. Синтез компонентов вируса.
Тема 1. Вирусная транскрипция.
Типы клеточных РНК-полимераз. Понятие о транскрипции вирусных ДНК. Этапы транскрипции. Механизмы инициации транскрипции. Регуляция белкового синтеза на уровне транскрипции. Кэпирование и поладенилирование. Сплайсинг пре-мРНК. Контроль процессинга мРНК при ретровирусной инфекции. Полиаденилирование и сплайсинг при аденовирусной инфекции. Регуляция альтернативного сплайсинга вирусными белками.
Тема 2. Обратная транскрипция.
История открытия и изучения процесса обратной транскрипции. Ретровирусы. Функции вирусной обратной транскриптазы. Механизмы интеграции. Провирус. Ретроэлементы в геноме эукариот. Вирус гепатита В. Строение частиц вируса гепатита В. Структура генома, продукты генов и их функции. Транскрипция ДНК, синтез прегеномной РНК. Механизм репликации вирусного генома, транслокация белковой и РНК-затравок. Общая схема жизненного цикла вируса гепатита В. Эпидемиология гепатита В и его роль в возникновении гепатокарцином.
Тема 3. Синтез вирусных белков.
Тема 4. Формирование вирусных частиц.
Общие принципы сборки вирусных частиц. Механизмы формирования зрелых вирусных частиц. Локализация белков в месте отпочковывания. Механизмы почкования вирусных частиц. Процессы отпочковывания на примере вируса гриппа и ВИЧ. Упаковка вирусной нуклеиновой кислоты. Сигналы упаковки ДНК-содержащих и РНК-содержащих вирусов, вирусов с сегментированным геномом.
Раздел IV. Онкогенные вирусы и медленные инфекции.
Тема 1. РНК-содержащие онкогенные вирусы.
Онкоген, протонкоген, анти-онкоген (супрессор опухоли) – определение понятий.
Онкогенные РНК-содержащие вирусы. Типы структурной организации геномов ретровирусов. Онкогенез в результате вставки промотора, энхансера, трансактивации клеточного генома (вирус Т-клеточного лейкоза человека).
Тема 2. ДНК-содержащие онкогенные вирусы.
Онкогенные ДНК-содержащие вирусы. Истинные вирусные онкогены (механизм действия). ДНК-содержащие онкогенные вирусы как модельная система для исследования молекулярной биологии эукариотической клетки.
Паповавирусы. Общая характеристика. Папилломавирусы. Полиомавирусы. Обезьяний вирус-40 (SV-40): структура, продуктивная и непродуктивная формы инфекции, литическая инфекция. Ранняя транскрипция генов SV-40. Т-антиген - мультифункциональный белок. Репликация ДНК SV-40. Поздняя транскрипция генов. Сборка вириона.
Тема 3. Медленные вирусные инфекции.
Характеристика медленных вирусных инфекций. Факультативные возбудители: вирус кори (подострый склерозирующий панэнцефалит), вирус краснухи (прогрессирующая врожденная краснуха, прогрессирующий краснушный панэнцефалит), вирус клещевого энцефалита (прогредиентная форма клещевого энцефалита), вирус простого герпеса (подострый герептический энцефалит), вирус бешенства (медленная рабическая инфекция). Облигатные возбудители: вирусы JC, ВК, ВИЧ, HTLV 1,2.
Тема 4. Прионные болезни.
Губчатые (спонгиформные) энцефалопатии, свойства, отличающие прионные болезни от других инфекционных заболеваний; история открытия. Роль белка РrРC в организме; Структура и свойства клеточного белка PrPc. Изменение характера укладки полипептидной цепи при превращении белка PrPC в прион PrPSc; механизм копирования конформации. Прионные заболевания человека: наследственные, инфекционные, спорадические; межвидовой барьер, диагностика. Сателлиты. Классификация сателлитов. Сателлитные вирусы. Сателлитные нуклеиновые кислоты (вирусоиды). Вирус гепатита дельта.
VIII. Перечень компетенций, формируемых в результате освоения дисциплины
В результате освоения дисциплины студент должен
· представлять себе место вирусов и субвирусных агентов в системе живой природы
· знать принципы строения вирусов и субвирусных агентов, устройство вирусных геномов вирусов различных групп и способы их экспрессии, способы взаимодействия вирусов и субвирусных агентов с другими организмами.
· уметь использовать полученные знания для применения в области фармации, медицины, медицинской, молекулярной, ветеринарной вирусологии, молекулярной и клеточной биологии, а также смежных областях.
IX. Используемые образовательные, научно-исследовательские и научно-производственные технологии:
В преподавании дисциплины используются демонстрационные компьютерные технологии.
Х. Учебно-методическое обеспечение самостоятельной работы студентов, оценочные средства контроля успеваемости и промежуточной аттестации:
Примеры тем для семинаров:
1. История открытия вирусов
2. Гипотезы происхождения вирусов
3. Гигантские вирусы (мимивирус, мегавирус, пандоравирус)
5. Онколитическая виротерапия
6. Вирус Зика: эпидемиология, пути передачи, лечение
7. Вирус Эбола: эпидемиология, профилактика, лечение
8. Современные препараты для профилактики и лечения ВИЧ-инфекции/СПИДа. Микробициды
9. Вирусные препараты для лечения, профилактики и диагностики инфекционных заболеваний
10. Возбудители медленных вирусных инфекций. Прионы. Вироиды
XI. Учебно-методическое и информационное обеспечение дисциплины
1. Репликация (от ДНК к ДНК)
2. Транскрипция (от ДНК к РНК)(
3. Трансляция (от РНК к белку)
4. Обратная транскрипция (от РНК к кДНК)
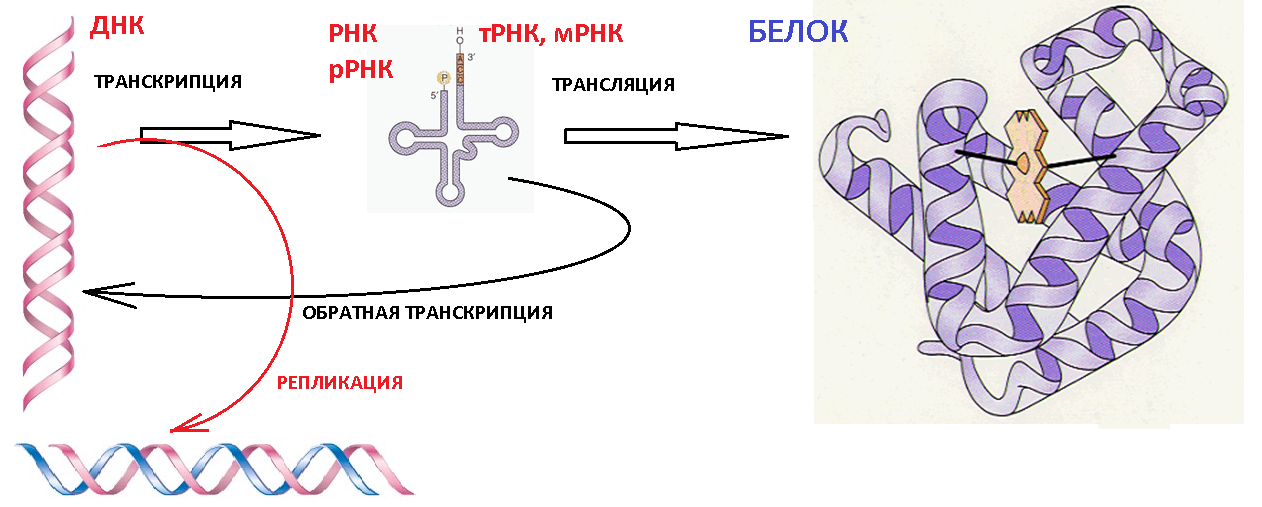
Долгое время считалось, что передача информации от РНК к ДНК невозможна, однако, впоследствии выяснилось, что это не так. Некоторые вирусы способны встраивать информацию со своей вирусной РНК в ДНК генома клетки-хозяина. Возможность "обратного" направления информации в настоящее время все шире используется в различных целях, от исследовательских до терапевтических. Так называемые энзимы- реверс-транскриптазы- способны осуществлять синтез кДНК на матрице РНК. О происходящих в клетках млекопитающих (эукариот) процессах передачи информации известно достаточно много, но далеко не все, и изложение хотя бы известных на данный момент времени сведений потребовалось бы слишком много места. Поэтому далее будут изложены лишь самые основы протекающих в клетках простейших организмов (прокариот) этапов передачи наследственной информации.
В процессе копирования информации происходит синтез дочерних молекул ДНК на основе информации, "записанной" в родительской молекуле ДНК. Ясно, что дочерние молекулы должны представлять собой точные копии родительской.
Репликация может осуществляться тремя способами:
а) консервативным;
б) полуконсервативным;
в) дисперсивным.
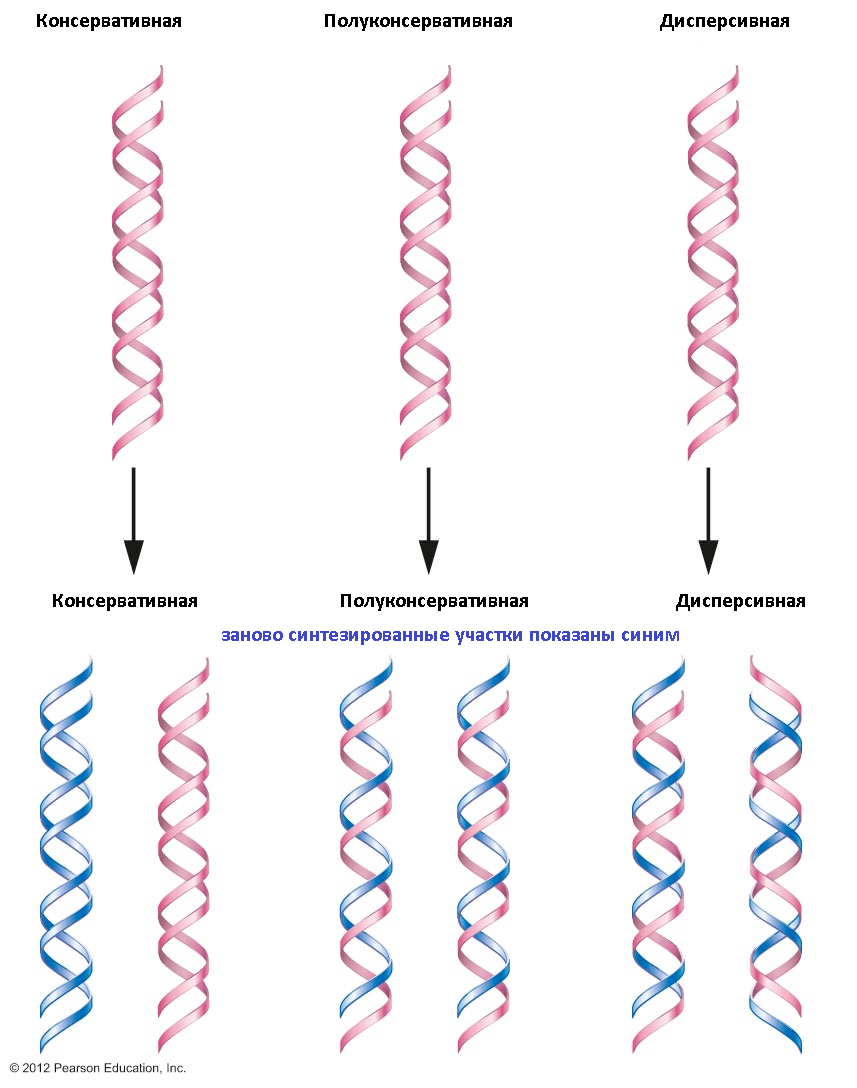
При консервативной репликации вновь синтезированные цепи ДНК находятся в дочерней молекуле. При полукон сервативной репликации полученные молекулы состоят из родительской и вновь синтезированной цепей. Дисперсивный способ репликации означает наличие перемежающихся родительских и вновь синтезированных участков на каждой из цепей образованных молекул ДНК. Для животных организмов и человека ХАРАКТЕРЕН только ПОЛУКОНСЕРВАТИВНЫЙ путь РЕПЛИКАЦИИ ДНК.
В процессе репликации участвует целый ряд энзимов (ферментов) с определенными функциями. Только синтезирующих ферментов в клетках прокариот насчитывается три. Их называют ДНК- полимеразами I, II и III. Сведения о функциональных особенностях ДНК-полимераз приведены в таблице.
| Проявляемая функция (активность) | |
| Полимеразная 5'-3' | |
| Экзонуклеазная 3'-5' | |
| Экзонуклеазная 5'-3' | |
| молекул в клетке | |
| производительность (нуклеотидов в мин, 37 С, на 1 молекулу Pol) |
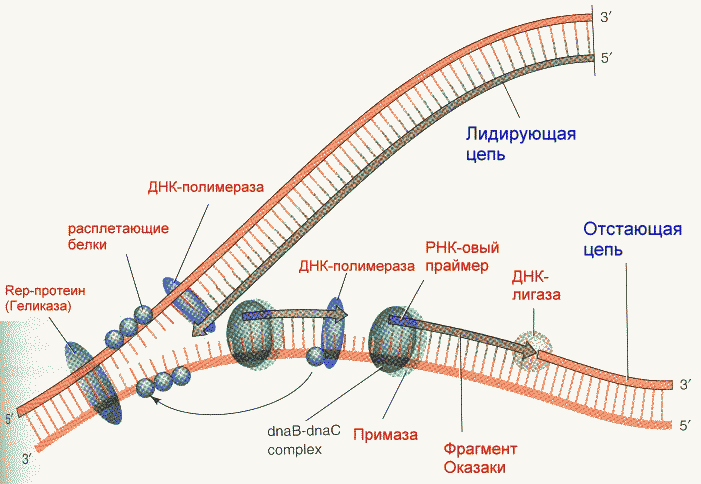
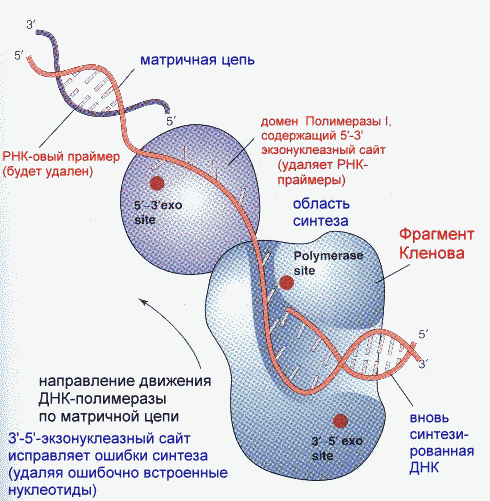
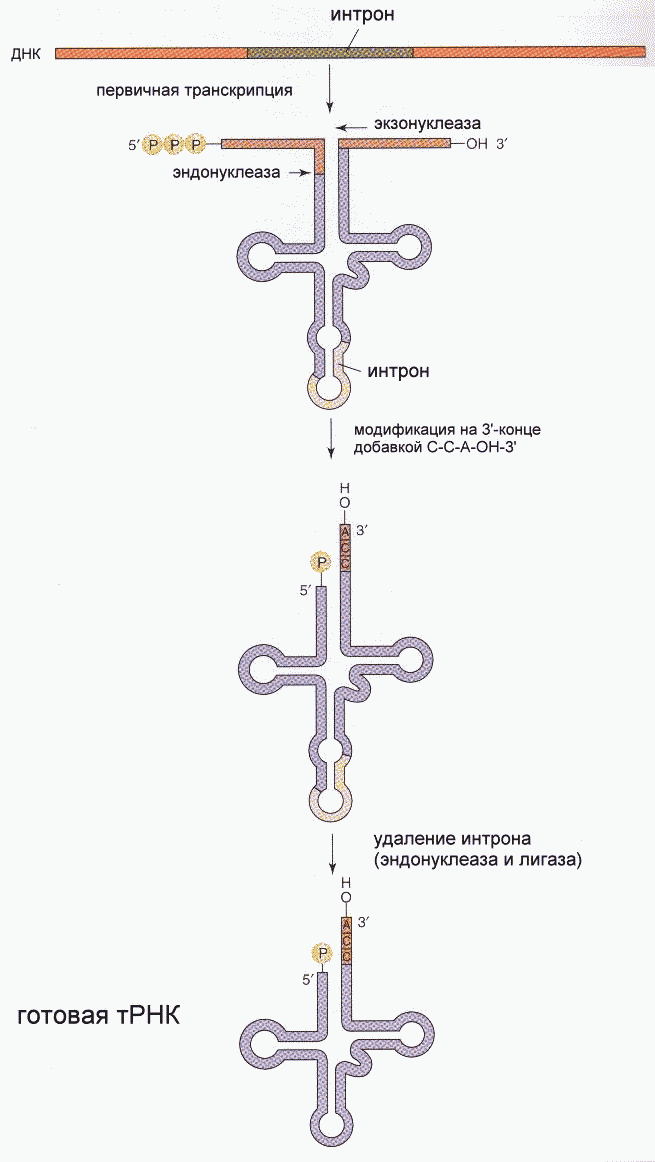
Фрагмент Кленова- результат частичного протеолиза ДНК-полимеразы I E. Coli субтилизином. Основная функция полимеразы III- синтез цепи, полимеразы I- синтез и исправление ошибочно вставленных нуклеотидов. Полимераза II осуществляет особые, спе циализированные функции. Репликация начинается с расплетания цепей ДНК специальными расплетающими белками, которые называют ГЕЛИКАЗАМИ (или Rep-протеином). Геликазы используют энергию АТФ в процессе расплетания цепей. Скорость расплетания составляет около 6000 мин-1. Для того, чтобы расплетенные цепи не могли вновь соединиться, имеются специальные SSB-белки (single-strand binding proteins), которые присоединяются к комплементарным цепям, удерживая их от ассоциации. По мере продвижения репликационной вилки SSB-протеины передвигаются по цепи, диссоциируя с одного места и присоединяясь на другом. Этот процесс не требует затрат энергии АТФ. После освобождения достаточного места начинается синтез праймера- затравки, необходимой для работы ДНК-полимеразы. Наличие затравки является необходимым условием функционирования ДНК-полимераз (как и наличие комплементарной цепи). В качестве затравки на каждой из разделенных цепей синтезируются маленькие отрезки молекул РНК при помощи фермента ПРИМАЗЫ. Синтез новой цепи ДНК осуществляется всегда в направлении 5'-3' , поэтому если по одной матричной цепи возможен непрерывный синтез, то по комплементарной ей цепи синтез осуществляется только участками. Эти участки синтеза называют фрагментами Оказаки. Когда синтез на одном из фрагментов Оказаки достигает праймера другого фрагмента, РНК-овый праймер удаляется имеющейся у полимераз 5'-3' экзонуклеазной активностью и достраивается дезоксирибонуклеотидами. После этого сахарофосфатный остов между фрагментами сшивается ковалентной связью при помощи фермента ДНК-лигазы. Частота возникновения ошибок при репликации и транскрипции НЕ ПРЕВЫШАЕТ 10 -8 -10 -9 , то есть возможна лишь одна ошибка на сотни миллионов нуклеотидов. Такая точность не может быть обеспечена одним только лишь правилом комплементарности нуклеотидов (обеспечивающим точность 1:10000-1:100000). Репликационный аппарат имеет собственные механизмы "поддержания точности" копирования генетической информации. Этими функциями обладают все ДНК-полимеразы. Модель структуры и функциональных участков (на примере ДНК-полимеразы I) показана на рисунке. Она имеет три зоны активности- полимеризующую в направлении 5'-3', и экозонуклеазные в направлениях 5'-3' и 3'-5'. Области активности разделены пространственно. Вперед (по ходу продвижения полимеразы по матричной цепи ДНК) обращена зона 5'-3' экзонуклеазной активности. Она служит для удаления попадающихся на пути РНК-овых праймеров (затравок). Далее идет собственно синтетическая зона и наконец, зона с экзонуклеазной активностью в направлении 3'-5'. С этой зоной связана так называемая PROOF-READING активность (способность узнавать неправильно встроенные нуклеотиды) и исправлять их вырезанием ряда уже встроенных нуклеотидов. Для этого молекула ДНК-полимеразы смещается (не отсоединяясь от ДНК-овой матрицы) к месту синтеза и последовательно вырезает нуклеотиды, после чего возобновляется нормальный синтез. Воздействие на организм неблагоприятных факторов (химические соединения, ультрафиолет и др.) приводит к постоянному накоплению ошибок в геноме, которые, в конечном итоге, вызывают появление патологии, в частности, невыясненный до сих пор механизм раковых заболеваний. Пока лишь существуют только предположения о том, что причиной раковых заболеваний являются дефекты в носителях информации- ДНК. Транскрипция- синтез молекул РНК на основании информации, записанной в ДНК. Осуществляется в ядрах при участии ДНК-зависимых РНК-полимераз, существующих в типах I, II и III (в порядке выхода в гель-хроматографии). В процессе транскрипции копируется не вся информация с ДНК, а только выборочная, часто отрезками. Сигналом для присоединения полимеразы служат так называемые промотеры , в районе которого (35 нуклеотидных пар до и 10 пар после него) и присоединяется РНК-полимераза. Происходит разделение цепей ДНК и начинается синтез молекулы РНК в направлении 5'-3', только на одной из цепей. При этом по месту тиминовых нуклеотидов комплементарной цепи встают уридиловые нуклеотиды. Весь комплекс передвигается по молекуле ДНК, пока не будет закончен синтез требуемого участка РНК. ДНК с "отсканированной" информацией репарирует, ассоциируя в двунитевые молекулы. Возбудитель туберкулеза Micobacterium tuberculosis (точнее, его РНК-полимераза) весьма чувствителен к антибиотику РИФАМПИЦИНУ, в то время как человеческая РНК-полимераза к нему мало чувствительна. На этом свойстве рифампицина основано его использование в терапии туберкулеза. Молекулы РНК очень часто претерпевают посттранскрипционную модификацию, заключающуюся в удалении участков построенной цепи. Наглядно это можно проследить на примере синтеза молекулы транспортной РНК: Читайте также:
 Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу. Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу.
Copyright © Иммунитет и инфекции
|