
Проникновение вируса в клетку виропексис характеристика
жЙЪЙПМПЗЙС ЧЙТХУПЧ 
чЙТХУ СЧМСЕФУС ПВМЙЗБФОЩН ЧОХФТЙЛМЕФПЮОЩН РБТБЪЙФПН Й ДМС ТБЪНОПЦЕОЙС ЕНХ ФТЕВХЕФУС ЦЙЧБС ЛМЕФЛБ. тБЪМЙЮБАФ ФТЙ ФЙРБ ЧЪБЙНПДЕКУФЧЙС ЧЙТХУБ У ЛМЕФЛПК:
- РТПДХЛФЙЧОЩК, ЙМЙ ГЙФПГЙДОЩК ФЙР, РТЙ ЛПФПТПН Ч ЪБТБЦЕООЩИ ЛМЕФЛБИ ПВТБЪХЕФУС ОПЧПЕ РПЛПМЕОЙЕ ЧЙТЙПОПЧ;
- БВПТФЙЧОЩК ФЙР, ИБТБЛФЕТЙЪХАЭЙКУС РТЕТЩЧБОЙЕН ЙОЖЕЛГЙПООПЗП РТПГЕУУБ Ч ЛМЕФЛЕ, РПЬФПНХ ОПЧЩЕ ЧЙТЙПОЩ ОЕ ПВТБЪХАФУС;
- ЙОФЕЗТБФЙЧОЩК ФЙР, ЙМЙ ЧЙТПЗЕОЙС, ЪБЛМАЮБАЭЙКУС Ч ЙОФЕЗТБГЙЙ, Ф.Е. ЧУФТБЙЧБОЙЙ ЧЙТХУОПК дол Ч ЧЙДЕ РТПЧЙТХУБ Ч ИТПНПУПНХ ЛМЕФЛЙ Й ЙИ УПЧНЕУФОПН УПУХЭЕУФЧПЧБОЙЙ.
рТПДХЛФЙЧОЩК ФЙР ЧЪБЙНПДЕКУФЧЙС ЧЙТХУБ У ЛМЕФЛПК ПУХЭЕУФЧМСЕФУС Ч ТЕЪХМШФБФЕ ТБЪНОПЦЕОЙС, Ф.Е. ТЕРТПДХЛГЙЙ ЧЙТХУБ (ПФ БОЗМ. reproduce - ЧПУРТПЙЪЧПДЙФШ). тЕРТПДХЛГЙС ЧЙТХУБ РТПЙУИПДЙФ Ч ОЕУЛПМШЛП УФБДЙК, ТБЪМЙЮБАЭЙИУС Х ТБЪОЩИ ЧЙТХУПЧ:
- БДУПТВГЙС ЧЙТЙПОПЧ ОБ ЛМЕФЛЕ;
- РТПОЙЛОПЧЕОЙЕ ЧЙТХУПЧ Ч ЛМЕФЛХ;
- ДЕРТПФЕЙОЙЪБГЙС ЙМЙ `ТБЪДЕЧБОЙЕ' ЧЙТХУПЧ Й ЧЩУЧПВПЦДЕОЙЕ ЧЙТХУОПЗП ЗЕОПНБ;
- ВЙПУЙОФЕЪ ЛПНРПОЕОФПЧ ЧЙТХУБ;
- ЖПТНЙТПЧБОЙЕ ЧЙТХУОПК ЮБУФЙГЩ;
- ЧЩИПД ЧЙТЙПОПЧ ЙЪ ЛМЕФЛЙ.
чЙТХУОПЕ ЙОЖЙГЙТПЧБОЙЕ ЛМЕФЛЙ ОБЮЙОБЕФУС У БДУПТВГЙЙ ЧЙТХУБ ОБ ЕЕ РПЧЕТИОПУФЙ (ТЙУ.3). бДУПТВГЙС ЧЙТХУБ ПВЕУРЕЮЙЧБЕФУС ЧЪБЙНПДЕКУФЧЙЕН ЕЗП РПЧЕТИОПУФОЩИ ВЕМЛПЧ УП УРЕГЙЖЙЮЕУЛЙНЙ ТЕГЕРФПТБНЙ ЮХЧУФЧЙФЕМШОЩИ ЛМЕФПЛ. уППФЧЕФУФЧЙЕ ЛМЕФПЮОЩИ ТЕГЕРФПТПЧ Й РПЧЕТИОПУФОЩИ ЧЙТХУОЩИ ВЕМЛПЧ ПРТЕДЕМСЕФ ФТПРЙЪН ЧЙТХУБ (ЗТЕЮ. tropos - РПЧПТПФ, ОБРТБЧМЕОЙЕ), ФП ЕУФШ УРПУПВОПУФШ ЙЪВЙТБФЕМШОП РПТБЦБФШ ПРТЕДЕМЕООЩЕ ЛМЕФЛЙ. чЙТХУЩ, ТЕРТПДХГЙТХАЭЙЕУС Ч ЛМЕФЛБИ РЕЮЕОЙ, ОБЪЩЧБАФУС ЗЕРБФПФТПРОЩНЙ, Ч ЛМЕФЛБИ ОЕТЧОПК УЙУФЕНЩ - ОЕКТПФТПРОЩНЙ Й Ф.Д.
рТПОЙЛОПЧЕОЙЕ ЧЙТХУБ Ч ЛМЕФЛХ РТПЙУИПДЙФ МЙВП РХФЕН ЧЙТПРЕЛУЙУБ (ТЕГЕРФПТОПЗП ЬОДПГЙФПЪБ), МЙВП УМЙСОЙС ПВПМПЮЛЙ ЧЙТХУБ У ЛМЕФПЮОПК НЕНВТБОПК (РТЙ ОБМЙЮЙЙ ВЕМЛБ УМЙСОЙС), ЙМЙ Ч ТЕЪХМШФБФЕ УПЮЕФБОЙС ЬФЙИ ДЧХИ НЕИБОЙЪНПЧ.
ч РТПГЕУУЕ РТПОЙЛОПЧЕОЙС ЧЙТЙПОБ Ч ЛМЕФЛХ РТЙ ХЮБУФЙЙ ЛМЕФПЮОЩИ ЖЕТНЕОФПЧ РТПЙУИПДЙФ ЕЗП ДЕРТПФЕЙОЙЪБГЙС, Ч ТЕЪХМШФБФЕ ЛПФПТПК ХДБМСАФУС РПЧЕТИОПУФОЩЕ УФТХЛФХТЩ ЧЙТХУБ, Й ЧЩУЧПВПЦДБЕФУС ЕЗП ЧОХФТЕООЙК ЛПНРПОЕОФ (УЕТДГЕЧЙОБ, ОХЛМЕПЛБРУЙД, ОХЛМЕЙОПЧБС ЛЙУМПФБ).
вЙПУЙОФЕЪ ЧЙТХУОЩИ ЛПНРПОЕОФПЧ ПУХЭЕУФЧМСЕФУС Ч ТБЪОЩИ ЮБУФСИ ЛМЕФЛЙ, РПЬФПНХ ОБЪЩЧБЕФУС ДЙЪЯАОЛФЙЧОЩН (ПФ МБФ. disjunctus - ТБЪПВЭЕООЩК). вЕМЛЙ ЧЙТХУБ УЙОФЕЪЙТХАФУС Ч ТЕЪХМШФБФЕ ФТБОУЛТЙРГЙЙ, Ф.Е. `РЕТЕРЙУЩЧБОЙС' ЙОЖПТНБГЙЙ У ЗЕОПНБ ЧЙТХУБ ОБ ЙОЖПТНБГЙПООХА тол (Йтол) Й РПУМЕДХАЭЕК ФТБОУМСГЙЙ (УЮЙФЩЧБОЙЕ Йтол ОБ ТЙВПУПНБИ) У ПВТБЪПЧБОЙЕН ВЕМЛБ ЧЙТХУБ. чЙТХУОБС ОХЛМЕЙОПЧБС ЛЙУМПФБ ЛПДЙТХЕФ УЙОФЕЪ УФТХЛФХТОЩИ Й ОЕУФТХЛФХТОЩИ ВЕМЛПЧ ЧЙТХУБ. уФТХЛФХТОЩЕ ВЕМЛЙ ЧИПДСФ Ч УПУФБЧ ЧЙТЙПОБ, Б ОЕУФТХЛФХТОЩЕ - СЧМСАФУС ЖЕТНЕОФБНЙ Й ПВЕУРЕЮЙЧБАФ ТЕРТПДХЛГЙА ЧЙТХУБ. пДОПЧТЕНЕООП У УЙОФЕЪПН ВЕМЛБ Ч ЛМЕФЛЕ РТПЙУИПДЙФ Й ТЕРМЙЛБГЙС (ПФ МБФ. replicatio - РПЧФПТЕОЙЕ), Ф.Е. УЙОФЕЪ ЧЙТХУОЩИ ОХЛМЕЙОПЧЩИ ЛЙУМПФ.
жПТНЙТПЧБОЙЕ ЧЙТЙПОПЧ РТПЙУИПДЙФ РХФЕН УБНПУВПТЛЙ: УПУФБЧОЩЕ ЮБУФЙ ЧЙТЙПОБ ФТБОУРПТФЙТХАФУС Ч НЕУФБ УВПТЛЙ ЧЙТХУБ Ч СДТЕ ЙМЙ ГЙФПРМБЪНЕ. уВПТЛБ ЛПНРПОЕОФПЧ ЧЙТЙПОБ РТПЙУИПДЙФ ЪБ УЮЕФ ЗЙДТПЖПВОЩИ, ЙПООЩИ, ЧПДПТПДОЩИ УЧСЪЕК Й УФЕТЙЮЕУЛПЗП УППФЧЕФУФЧЙС. ч ТЕЪХМШФБФЕ УБНПУВПТЛЙ ЛБРУПНЕТПЧ, ПВТБЪПЧБЧЫЙИУС ЙЪ ЧЙТХУОЩИ РПМЙРЕРФЙДПЧ, Й ЧЪБЙНПДЕКУФЧЙС ЙИ У ОХЛМЕЙОПЧЩНЙ ЛЙУМПФБНЙ ЧЙТХУБ ПВТБЪХАФУС ОХЛМЕПЛБРУЙДЩ. уХРЕТЛБРУЙДОБС ПВПМПЮЛБ УМПЦОППТЗБОЙЪПЧБООЩИ ЧЙТХУПЧ ЧЛМАЮБЕФ Ч УЕВС ЛТПНЕ ЧЙТХУУРЕГЙЖЙЮЕУЛЙИ ВЕМЛПЧ ЕЭЕ ЛПНРПОЕОФЩ НЕНВТБОЩ ЛМЕФЛЙ.
тЙУ.3. уФБДЙЙ ТЕРТПДХЛГЙЙ ЧЙТХУПЧ.
1 - БДУПТВГЙС ЧЙТЙПОБ ОБ ЛМЕФЛЕ; 2 - РТПОЙЛОПЧЕОЙЕ ЧЙТЙПОБ Ч ЛМЕФЛХ РХФЕН ЧЙТПРЕЛУЙУБ;
3 - ЧЙТХУ ЧОХФТЙ ЧБЛХПМЙ ЛМЕФЛЙ; 4 - `ТБЪДЕЧБОЙЕ ЧЙТЙПОБ ЧЙТХУБ; 5 - ТЕРМЙЛБГЙС ЧЙТХУОПК ОХЛМЕЙОПЧПК ЛЙУМПФЩ; 6 - УЙОФЕЪ ЧЙТХУОЩИ ВЕМЛПЧ ОБ ТЙВПУПНБИ ЛМЕФЛЙ; 7 - ЖПТНЙТПЧБОЙЕ ЧЙТЙПОБ; 8 - ЧЩИПД ЧЙТЙПОБ ЙЪ ЛМЕФЛЙ РХФЕН РПЮЛПЧБОЙС. (нЙЛТПВЙПМПЗЙС Й ЙННХОПМПЗЙС рПД ТЕДБЛГЙЕК чПТПВШЕЧБ б.б. - н. - 1999)
Типы взаимодействия вируса с клеткой. Различают три типа взаимодействия вируса с клеткой: продуктивный, абортивный и ин-тегративный.
Продуктивный тип — завершается образованием нового поколения вирионов и гибелью (лизисом) зараженных клеток (цитоли-тическая форма). Некоторые вирусы выходят из клеток, не разрушая их (нецитолитическая форма).
Абортивный тип — не завершается образованием новых вирионов, поскольку инфекционный процесс в клетке прерывается на одном из этапов.
Интегративный тип, или вирогения — характеризуется встраиванием (интеграцией) вирусной ДНК в виде провируса в хромосому клетки и их совместным сосуществованием (совместная репликация).
Адсорбция. Взаимодействие вируса с клеткой начинается с процесса адсорбции, т. е. прикрепления вирусов к поверхности клетки. Это высокоспецифический процесс. Вирус адсорбируется на определенных участках клеточной мембраны — так называемых рецепторах. Клеточные рецепторы могут иметь разную химическую природу, представляя собой белки, углеводные компоненты белков и липидов, липиды. Число специфических рецепторов на поверхности одной клетки колеблется от 10 4 до 10 5 . Следовательно, на клетке могут адсорбироваться десятки и даже сотни вирусных частиц.
Проникновение в клетку. Существует два способа проникновения вирусов животных в клетку: виропексис и слияние вирусной оболочки с клеточной мембраной. При виропексисе после адсорбции вирусов происходят инвагинация (впячивание) участка клеточной мембраны и образование внутриклеточной вакуоли, которая содержит вирусную частицу. Вакуоль с вирусом может транспортироваться в любом направлении в разные участки цитоплазмы или ядро клетки. Процесс слияния осуществляется одним из поверхностных вирусных белков капсидной или суперкапсидной оболочки. По-видимому, оба механизма проникновения вируса в клетку не исключают, а дополняют друг друга.
Биосинтез компонентов вируса. Проникшая в клетку вирусная нуклеиновая кислота несет генетическую информацию, которая успешно конкурирует с генетической информацией клетки. Она дезорганизует работу клеточных систем, подавляет собственный метаболизм клетки и заставляет ее синтезировать новые вирусные белки и нуклеиновые кислоты, идущие на построение вирусного потомства.
Реализация генетической информации вируса осуществляется в соответствии с процессами транскрипции, трансляции и репликации.
Существуют следующие общие принципы сборки вирусов, имеющих разную структуру:
1. Формирование вирусов является многоступенчатым процессом с образованием промежуточных форм;
2. Сборка просто устроенных вирусов заключается во взаимодействии молекул вирусных нуклеиновых кислот с капсидными белками и образовании нуклеокапсидов (например, вирусы полиомиелита). У сложно устроенных вирусов сначала формируются нуклеокапсиды, с которыми взаимодействуют белки суперкапсидных оболочек (например, вирусы гриппа);
3. Формирование вирусов происходит не во внутриклеточной жидкости, а на ядерных или цитоплазматических мембранах клетки;
4. Сложно организованные вирусы в процессе формирования включают в свой состав компоненты клетки-хозяина (липиды, углеводы).
Время, необходимое для осуществления полного цикла репродукции вирусов, варьирует от 5—6 ч (вирусы гриппа, натуральной оспы и др.) до нескольких суток (вирусы кори, аденовирусы и др.). Образовавшиеся вирусы способны инфицировать новые клетки и проходить в них указанный выше цикл репродукции. Взаимодействие вируса с клеткой хозяина.
Существует несколько типов взаимодействия с клеткой.
Жизненный цикл вирусов:
1. Продуктивная инфекция протекает в несколько стадий:
1. Адсорбция вириона на клетке – осуществляется за счёт специфического взаимодействия антирецептора вириона с комплементарным рецепторами мембраны клетки хозяина. Не зависит от температуры.
2. Проникновение вируса в цитоплазму клетки (пенетрация). У просто устроенных вирусов осуществляется путем эндоцитоза – образуется эндоцитарная вакуоль, в которую заключен вирион. У сложноустроенных вирусов осуществляется путем слияния клеточной мембраны с мембраной суперкапсида.
3. Депротеинизация (раздевание вируса). Осуществляется клеточными ферментами, разрушающими капсид. При этом вирусный геном освобождается от капсида (белка). Вирус на время как бы исчезает. Эта стадия называется эклипс.
4. Синтез вирусных компонентов (генома и белка). Синтез разобщен во времени и протекает в разных частях клетки. На этой стадии синтезируются вирусные белки и нуклеиновые кислоты.
Стратегия вирусного генома
ДНК - содержащие вирусы: аденовирусы, ретровирусы (онкогенны), вирус гепатита В, герпес;
• В ядре пораженной клетки с вирусной ДНК синтезируется информационная РНК с учатием ДНК-полимеразы. Днк – иРНК – ДНК вируса.
• У вирусов с +РНК пикорна-, тогавирусы., гепатит А, Е и др., функцию иРНК выполняет сам геном. Все компоненты синтезируются только в цитоплазме. РНК вируса - +РНК – белок вируса.
• У вирусов с –РНК орто-, парамиксо-, рабдовирусы и др. В цитоплазме пораженной клетки. РНК вируса – РНК – иРНК – белок вируса.
• РНК-содержащие ретровирусы (ВИЧ) имеют обратную транскриптазу или ревертазу. В ядре пораженной клетки с РНК синтезируется ДНК вируса. РНК вируса – РНК – ДНК – иРНК – белок вируса
Ферменты, осуществляющие синтез вирусных нуклеиновых кислот:
• РНК-зависимая – ДНК-полимераза (обратная транскриптаза).
5. Сборка вирионов
• Может происходить в цитоплазме или на внутренней поверхности клеточной мембраны.
• Осуществляется по принципу самосборки на основе белок-белкового и белок-нуклеинового узнавания.
6. Выход из клетки.
• Просто устроенные вирионы выходят из клетки путем взрыва;
• Сложно устроенные выходят методом почкования.
2. Стадия интеграции
У вирусов герпеса, гепатита В, ВИЧ в цикле репродукции имеется стадия интеграции (встраивания) вирусного генома в геном клетки хозяина.
Интегрированная в клеточный геном вирусная ДНК называется провирусом.
!Вирусы не растут на питательных средах!. Для культивирования вирусов используются биологические модели:
Вирус (от лат. virus — яд) — простейшая форма жизни, микроскопическая частица, представляющая собой молекулы нуклеиновых кислот (ДНК или РНК, некоторые, например, мимивирусы, имеют оба типа молекул), заключённые в белковую оболочку и способные инфицировать живые организмы.
От других инфекционных агентов вирусы отличает капсид (белковая оболочка). Вирусы, за редким исключением, содержат только один тип нуклеиновой кислоты: либо ДНК, либо РНК. Ранее к вирусам также ошибочно относили прионы, однако впоследствии оказалось, что эти возбудители представляют собой особые белки и не содержат нуклеиновых кислот.
Вирусы являются облигатными паразитами, так как вирусы не способны размножаться вне клетки. Вне клетки вирусные частицы ведут себя как химические вещества. В настоящее время известны вирусы, размножающиеся в клетках растений, животных, грибов и бактерий (последних обычно называют бактериофагами). Обнаружены также вирусы, поражающие другие вирусы (вирусы-сателлиты).
Строение и свойства
Размеры вирусов колеблются от 20 до 300 нм. В среднем они в 50 раз меньше бактерий. Их нельзя увидеть в световой микроскоп, так как их длины меньше длины световой волны.
Вирусы состоят из различных компонентов:
- сердцевина / генетический материал (ДНК или РНК).
- белковая оболочка, которую называют капсидом.
- дополнительная липопротеидная оболочка.
Полностью сформированная инфекционная частица называется вирионом.
Генетический аппарат вируса несет информацию о нескольких типах белков, которые необходимы для образования нового вируса: ген, кодирующий обратную транскриптазу и другие.
Капсид — это внешняя оболочка вируса, состоящая из белков. Капсид выполняет несколько функций.
- Защита генетического материала (ДНК или РНК) вируса от механических и химических повреждений.
- Определение потенциала к заражению клетки.
- На начальных стадиях заражения клетки: прикрепление к клеточной мембране, разрыв мембраны и внедрение в клетку генетического материала вируса.
Дополнительная липопротеидная оболочка образована из плазматической мембраны клетки-хозяина. Она встречается только у сравнительно больших вирусов (грипп, герпес). Эта наружная оболочка является фрагментом ядерной или цитоплазматической мембраны клетки-хозяина, из которой вирус выходит во внеклеточную среду. Иногда в наружных оболочках сложных вирусов помимо белков содержатся углеводы, например у возбудителей гриппа и герпеса.
Каждый компонент вирионов имеет определённые функции: белковая оболочка защищает их от неблагоприятных воздействий, нуклеиновая кислота отвечает за наследственные и инфекционные свойства и играет ведущую роль в изменчивости вирусов, а ферменты участвуют в их размножении.
Более сложные по структуре вирусы, кроме белков и нуклеиновых кислот, содержат углеводы, липиды. Для каждой группы вирусов характерен свой набор белков, жиров, углеводов и нуклеиновых кислот. Некоторые вирусы содержат в своём составе ферменты.
Взаимодействие вируса с клеткой
Вирусы вне клетки представляют собой кристаллы, но при попадании в клетку “оживают”. Их размножение происходит особым, ни с чем не сравнимым способом. Сначала вирионы проникают внутрь клетки, и освобождаются вирусные нуклеиновые кислоты. Затем "заготавливаются" детали будущих вирионов. Размножение заканчивается сборкой новых вирионов и выходом их в окружающую среду.
Встреча вирусов с клетками начинается с его адсорбции, то есть прикрепления к клеточной стенке. Затем начинается внедрение или проникновение вириона в клетку, которое осуществляет она сама. Однако, как правило, проникновению вируса в цитоплазму клетки предшествует связывание его с особым белком-рецептором, находящимся на клеточной поверхности.
Связывание с рецептором осуществляется благодаря наличию специальных белков на поверхности вирусной частицы, которые "узнают" соответствующий рецептор на поверхности чувствительной клетки. На одной клетке могут адсорбироваться десятки, и даже сотни вирионов. Участок поверхности клетки, к которому присоединился вирус, погружается в клетку. Этот процесс называется виропексисом.
Инфекционный процесс начинается, когда проникшие в клетку вирусы начинают размножаться, т.е. происходит редупликация вирусного генома и самосборка капсида. Для осуществления редупликации нуклеиновая кислота должна освободиться от капсида. После синтеза новой молекулы нуклеиновой кислоты она одевается, синтезированными в цитоплазме клетки – вирусными белками – образуется капсид.
Накопление вирусных частиц приводит к выходу их из клетки. Для некоторых вирусов это происходит путем "взрыва", в результате чего целостность клетки нарушается и она погибает другие вирусы выделяются способом, напоминающим почкование. В этом случае клетки организма могут долго сохранять свою жизнеспособность.
Поразительно, как вирусы, которые в десятки и даже сотни раз меньше клеток, умело, и уверенно распоряжаются клеточным хозяйством. Размножаясь, они истощают клеточные ресурсы и глубоко, часто необратимо, нарушают обмен веществ, что, в конечном счете, является причиной гибели клеток.
Типы вирусных инфекций
В зависимости от длительности пребывания вируса в клетке и характера изменения её функционирования различают три типа вирусной инфекции.
Если образующиеся вирусы одновременно покидают клетку, то она разрывается и гибнет. Вышедшие из неё вирусы поражают новые клетки. Так развивается литическая (разрушение, растворение) инфекция.
При вирусной инфекции другого типа, называемой персистентной (стойкой), новые вирусы покидают клетку-хозяина постепенно. Клетка продолжает жить и делится, производя новые вирусы, хотя её функционирование может измениться.
Третий тип инфекции называется латентным (скрытым). Генетический материал вируса встраивается в хромосомы клетки и при её делении воспроизводится и передаётся дочерними клетками.
Классификация
- По сердцевине: ДНК-содержащие и РНК-содержащие (ретро) вирусы.
- По структуре капсомеров: Изометрические (кубические), спиральные, смешанные.
- По наличию или отсутствию дополнительной липопротеидной оболочки:
- По клеткам-хозяинам.
Кроме этих классификаций есть еще много других. Например, по типу переноса инфекции от одного организма к другому.
Классификация и свойства вирусов гепатита:
| Семейство | Геном | Механизмы и пути передачи | Инфекция | ||
| острая | хроническая | ||||
| Вирус гепатита A (HAV) | Picornaviridae | РНК | Фекально-оральный | Да | Нет |
| Вирус гепатита B (HBV) | Hepadnaviridae | ДНК | Парентеральный, половой, перинатальный | Да | Да |
| Вирус гепатита C (HCV) | Flaviviridae | РНК | Парентеральный, половой, перинатальный | Да | Да |
| Вирус гепатита D (HDV) | Viroid | РНК | Парентеральный, половой, перинатальный | Да | Да |
| Вирус гепатита Е (HEV) | Caliciviridae | РНК | Фекально-оральный | Да | Нет |
| Вирус гепатита G (HGV) | Flaviviridae? | РНК | Перинатальный, трансмиссивный? | Да | Да |
| Вирус гепатита F (HFV) | мутант HBV? | ДНК? | Перинатальный, трансмиссивный? | Да | Да? |
Вирусные антигены
Антигены (АГ) - генетически чужеродные вещества, способные вызывать образование антител и эффекторов клеточного иммунитета. Иммунные реакции, вызванные антигенами, направлены на их связывание, нейтрализацию и выведение из организма.
Способность антигена вызывать устойчивость против заражения называется протективной активностью. Она является главной характеристикой антигенов любой вакцины.
Вирусные антигены могут входить в состав самого вириона или быть вирус-индуцированными и находиться в зараженной клетке. Антигены вируса могут располагаться на поверхности клетки. Это вирусные гликопротеиды, образующиеся в результате встраивания вируса в клеточную мембрану, репродукции оболочечного вируса или выхода вируса из клетки. Для некоторых вирусов характерна ярко выраженная изменчивость антигенной структуры.
Грубо говоря, вирусные антигены - это отдельные частички вируса или продукты его жизнедеятельности, которые вызывают срабатывание защитной реакции иммунной системы - выработку соответствующих антител.
Антитела (иммуноглобулины, ИГ, Ig) - это растворимые гликопротеины, присутствующие в сыворотке крови, тканевой жидкости или на клеточной мембране, которые распознают и связывают антигены. Антитела синтезируются иммунной системой в ответ на чужеродные вещества - антигены.
Все вирусные антигены имеют белковую природу; среди них - гликопротеины (обычно - поверхностные), фосфопротеины, нуклеопротеины. Чаще всего протективными являются поверхностные в вирионе гликопротеины, хотя образуемые в ходе иммунного ответа антитела направлены против многих белков, в том числе и расположенных в нуклеокапсиде, "в глубине" вириона.
Принципиальная, отличительная от других возбудителей особенность репродукции вирусов заключается в том, что не все белки, синтез которых индуцируется в инфицированной клетке, входят затем в состав вириона. Часть из них является вспомогательными, обеспечивающими процесс репродукции. Тем не менее, они также могут попадать во внеклеточную среду и служить иммунизирующим материалом.
У большинства вирусов имеются суперокапсидные - поверхностные оболочечные, белковые и гликопротеидные АГ (например, гемагглютинин и нейраминидаза вируса гриппа), капсидные - оболочечные и нуклеопротеидные (сердцевинные) АГ.
Вакцины (от лат. vacca – корова) – препараты микробного и вирусного происхождения, использующиеся для выработки в организме активного искусственного иммунитета. Вакцины готовят из живых аттенуированных микроорганизмов (живые вакцины), убитых микробов (убитые вакцины) или путем извлечения из бактериальных клеток антигенных комплексов (химические вакцины). См. Аттенуация. Термин впервые использован Дженнером в 1798 г. и распространен Пастером на все микробные препараты, способствующие выработке активного иммунитета.
Вегетативное размножение фага – внутриклеточное развитие фага, характеризующееся редупликацией (см.) фаговой ДНК и синтезом фагового белка. В это время отсутствуют зрелые, инфекционные частицы фага, а незрелые частицы не могут быть обнаружены путем подсчета стерильных пятен (см. Бляшки вирусов). См. Фаг вегетативный.
Версен (трилон Б) – натриевая соль этилендиаминтетрауксусной кислоты. При взаимодействии версена с поливалентными ионами образуются хелаты, представляющие собой внутрикомплексные соединения с солевой связью. В вирусологии применяется для отделения слоя клеток культуры ткани (см.) от стеклянной стенки сосуда при получении субкультур.
Вибрионы – бактерии, имеющие извитую форму (один завиток не более 1/4 оборота спирали). Размножаются поперечным делением.
Вирион (син. Вироспора) – единичная особь, отдельная вирусная частица, физическая единица какого-либо вируса. Представляет собой заключительную форму онтогенетического развития вируса. Термин введен Львовым, Андерсоном и Жакобом.
Вирогения – форма сосуществования , вируса с клеткой хозяина, при которой геном (см.) вируса включается в хромосому (см.) клетки (см. Провирус). При этом в некоторых случаях изменяются наследственные свойства клетки (см. Трансформация вирусная). Вирусы, вызывающие вирогению, называются умеренными. К ним можно отнести онкогенные вирусы. См. Лизогения. Вирусы онкогенные.
Виропексис – см. Пиноцитоз.
Вироспора (син. Вирион) – термин предложен Рыжковым в 1955 г. См. Вирион.
Вирулентность (от лат. virulentus – ядовитый, заразный) – количественная характеристика степени патогенности (см.) микроорганизма. Механизм вирулентности микроба связан с его способностью противостоять защитным реакциям макроорганизма или подавлять их факторами агрессивности и инвазивности. Агрессивность связана с химическими соединениями, входящими в состав поверхностных структур, бактерий (см. Капсула, Клеточная стенка). Изменения поверхностных структур (например, при переходе S-формы кишечных бактерий в R-форму, при утрате капсулы у пневмококков) сопровождаются потерей или ослаблением вирулентности микробов (см. Колонии гладкий S-тип, колонии шероховатый R-тип). Инвазивность определяется энзимами микроба, нарушающими целостность некоторых тканей макроорганизма. К ним относят фактор проницаемости (см. Гиалуронидаза), лецитиназу, коллагеназу, плазмокоагулазу (см.) и др. Отдельные химические компоненты бактериальной клетки сами по себе не обладают вирулентными свойствами. Вирулентность присуща только живой функционирующей микробной клетке, размножающейся в организме хозяина. Изложенное дает возможность считать вирулентность фенотипическим проявлением патогенного генотипа микроба. См. Генотип, Фенотип.
Вирус маскированный (скрытый, латентный) – форма существования вируса в клетке хозяина, которая не обнаруживается прямыми вирусологическими методами.
Вирус минимальный – простоорганизованный вирус или минимально необходимая для инфицирования часть полноценного вириона, состоящая из нуклеиновой кислоты и белковых капсомеров).
Вирус неполный (син. Неполноценный, Дефективный) – вирусная частица, лишенная некоторых фрагментов нуклеиновой кислоты. Дефектный вирус дает полноценное потомство, только если вместе с ним в клетке присутствует вирус-помощник (см.). См. Фаг дефективный.
Вирус помощник – вирус, геном которого способен восполнить путем комплементации (см.) ту метаболическую или морфогенетическую функцию, которая отсутствует у дефектного вирусного генома (см. Вирус неполный). Это достигается при совместном заражении обоими вирусами клетки хозяина или при заражении клетки хозяина, несущей дефектный вирус, вирусом-помощником.
Вируса филаментозные формы (от лат. fillum – нить) – нитевидные формы некоторых вирусов.
Вируса чистые линии – потомство, полученное из одной вирусной частицы. С этой целью в агаровом покрытии получают бляшки вирусов (см.). Одну из них рассевают для получения следующей генерации (см), из которой повторно отбирают одну бляшку и т. д. – до получения 5-10 генераций.
Вирусемия – проникновение вируса в кровь и распространение его с током крови.
Вирусная конверсия – см. Лизогенная конверсия.
Вирусные внутриклеточные включения – морфологические образования в цитоплазме или ядре клеток, пораженных вирусом. Могут быть выявлены при помощи специальных методов окраски. Различают следующие типы включений: 1) кристаллообразные скопления вирусов, расположенные в цитоплазме (вирус полиомиелита) или в ядре (аденовирус); 2) продукты реакции клетки на вирусную инфекцию (тельца при герметической инфекции); 3) сочетание скопления вирусов с продуктами реакции клетки (Бабеша-Негри тельца (см.) при бешенстве).
Вирусоносительство латентное – содержание вируса в тканевой культуре (см.) или в организме животных и человека без видимого проявления цитопатического действия (см.) или клинических симптомов заболевания. Латентное вирусоносительство описано у вирусов группы герпеса (см.), аденовирусов (см), энтеровирусов (см.), онкогенных вирусов (см.). Онкогенные вирусы могут встраиваться в хромосому клетки-хозяина, вызывая состояние, аналогичное лизогенизации бактерий (см. Вирогения, Лизогения).
Вирусы онкогенные – вирусы, индуцирующие образование опухолей у человека и животных, а также способные вызывать злокачественную трансформацию клеток (см.) в культурах ткани (см.). Выделено несколько десятков вирусов, вызывающих опухоли и лейкозы у амфибий, птиц, млекопитающих, в том числе и у человека (папилломы, кандиломы). Известны вирусные опухоли беспозвоночных животных и растений. Онкогенные вирусы могут содержать ДНК (см. Папова вирусы, Аденовирусы и др.), а также РНК (вирусы лейкозов кур и мышей, см. Онкорна-вирусы). Механизм онкогенного действия вирусов окончательно не выяснен. Для большинства из них характерно поведение вирусов интрагеномных (см.) и латентное вирусоносительство (см.). Для воспроизведения некоторых онкогенных вирусов в клетках обходимо присутствие вируса-помощника (см.). См. также Вирогения.
Вирусы экстрагеномные – вирусы, нуклеиновая кислота которых реплицируется автономно в чувствительных клетках. К ним относятся большинство известных вирусов животных, растений и бактерий (см. Фаги). Выделяются в противоположность, вирусам интрагеномным (см).
Витамины – незаменимые и ограниченно заменимые ростовые факторы, которые используют микробы при синтезе ряда коферментов и в качестве кофакторов (см.) в ряде метаболических реакций. По потребности в витаминах различные микроорганизмы отличаются друг от друга. К витаминам, в которых чаще всего нуждаются ауксотрофы (см.), относятся следующие: тиамин (В1), рибофлавин (В2), пантотеновая кислота (В3), никотиновая кислота (В5 или РР), пиридоксаль (В6), биотин (Н), липоевая кислота, фолиевая кислота (ВС), п-аминобензойная кислота, кобамиды (В12), железопорфирины и хиноны (К). Биологическая роль витаминов в механизме микробных клеток велика и разнообразна. Так, рибофлавин входит в состав флавопротеидов, участвующих в биологическом окислении; пантотеновая кислота входит в состав коэнзима А. Никотиновая кислота – в состав НАД (никотинамиддинуклеотида); пиридоксаль – в состав ферментов аминокислотного обмена. Способность микроорганизмов, обитающих в толстом кишечнике человека синтезировать ряд витаминов группы В (например В12) имеет большое значение для витаминного баланса макроорганизма.
Включения цитоплазмы у микроорганизмов – гранулы, непостоянно встречающиеся в цитоплазме микробных клеток. Чаще всего являются продуктами внутриклеточного метаболизма и служат запасными источниками питательных веществ. В микробных клетках встречаются включения волютина (см.), липоидных и жировых веществ, гликогена, полисахаридов, серы, железа и других соединений. Обычно включения разных веществ обнаруживают путем специальных окрасок или реакций.
Внехромосомные факторы наследственности – см. Эписомы.
Волютин, зерна волютина – включения, содержащиеся в цитоплазме некоторых бактерий (спирилл, дифтерийной палочки, дрожжей). Представляют собой комплекс РНК и полиметафосфата. По-видимому, выполняют роль резервных источников этих соединений или их компонентов. Существенная особенность зерен волютина – способность к метахромазии (изменению цвета красителя при окрашивании). Природа этого феномена заключается в том, что при сорбции некоторых красивей, на сильно заряженных полимерах (полифосфатах, нуклеиновых кислотах) происходит их полимеризация, сопровождающаяся изменением цвета. Зерна волютина окрашиваются уксуснокислой синькой в темно-синий цвет по способу Нейссера (см.).
Вирусы являются внутриклеточными инфекционными агентами. Весь репликативный цикл вируса осуществляется с исполь зованием метаболических и генетических ресурсов клеток. Поэтому патогенез вирусных инфекций, в первую очередь, следует рассматривать на молекулярном и клеточном уровнях (В.И. Покровский, О.И. Киселев, 2002). Вместе с тем, инфекционный процесс, вызванный вирусами, развивается в пределах того или иного органа или ткани, так как большинство вирусов обладают достаточно высокой органной или тканевой тропностью. Поэтому характер развития внутритканевых процес сов при вирусных инфекциях, с одной стороны, определяется, как правило, цитопатическим действием вируса на клетки данной ткани и органа, а с другой стороны, реакцией внутритканевых и органных систем защиты от вирусной инфекции. Последние не редко носят деструктивный характер и усугубляют течение забо левания и его последствия. Поэтому понимание патогенеза вирусных инфекций представляется исключительно важным для создания рациональных схем лечения, основанных на гармоничном сочетании симптоматического лечения и противовирусной химиотерапии.
Стадии репликативного цикла вируса гриппа представлены на рис. 1. Такой же репликативный цикл свойствен большинству РНК- и ДНК-содержащих вирусов, кроме тех, кото рые обладают особыми механизмами хронизации процесса, латенции и способностью к интеграции в клеточный геном (например, ВИЧ, гепатит С, вирус папилломы и.т.). Ключевыми стадиями репликативного цикла вируса являются проникновение в клетки, декапсидация, активация процессов транскрипции и трансляции, собственно репликация вирусного генома и созревание зрелых вирусных частиц с выходом нового потомства инфекционного вируса.
Проникновение вируса в клетки - одна из ключевых стадий патогенеза инфекционного процесса. Для большинства вирусов степень цитопатического действия вируса прямо зависит от множественности заражения. В естественных условиях инфицирова ние клеток вирусами не происходит обычно с высокой множе ственностью. Исключение составляют респираторные вирусы, которые передаются воздушно-капельным путем с высокой инфицирующей дозой. ВИЧ и гепатиты, наоборот, эффективно передаются с кровью в самых незначительных трудно определяе мых титрах. Такие вирусы имеют особые свойства, определяющие их контагиозность.
Для проникновения вируса в клетку, необходимо: высокое сродство к вирусоспецифическому рецептору; множественность рецепторов; эффективность слияния вируса с клетками при инфицировании; способность к образованию синцития, к образованию гигантских синпластов, состоящих из множества слившихся клеток, в которых вирусный нуклеопротеид без эндоцитоза беспрепятственно переходит от одной зараженной клетки к множеству других через цитоплазму.
Такими свойствами из респираторных вирусов обладает респираторно-синцитиальный вирус, а среди ретровирусов – вирус иммунодефицита человека.
Стадии жизненного цикла вируса гриппа и соответствующие ингибиторы репродукции в известной степени могут быть экстраполированы на многие другие вирусы, за исключением тех, жизненный цикл которых включает интеграцию в клеточный геном. В процессе репликации вирусов обычно происходит селективная блокада трансляции клеточных мРНК и активация экспресси клеточных генов, кодирующих провоспапительные лимфокины. Многократные раунды репликации вирусного генома приводят к истощению энер гетических ресурсов клеток и пула предшественников азотистых оснований. Повышенное содержание провоспалительных лимфокинов приводит к развитию воспалительной реакции и локально му генерированию свободных радикалов, оказывающих, в свою очередь, повреждающее действие на клетки и ткани через перекисное окисление липидов клеточных мембран, инактивацию жизненно важных ферментов. Поэтому одним из важнейших факторов патогенности вирусов является их репликативный потенциал: чем активнее происходит репликация, тем сильнее цитопатическое действие вирусов на клетки и пораженную ткань (орган).
Значительный вклад в цитопатическое действие вирусов вносит репродукция вирусов в митохондриях, что сопровождается нарушением функций этих органоидов, падением продукции АТФ и индукцией опосредованного митохондриями пути апоптоза.
Впервые возможность репродукции вирусов в митохондриях была доказана выдающимся отечественным вирусологом В.М. Ждановым. Узкий тропизм вирусов, в известной степени, спасителен для организма, так как инфекционный процесс ограничен одним органом.
Это обстоятельство позволяет, проводит рациональную симптоматическую терапию, направленную на защиту данной ткани или органа и ориентировать противовирусные средства в данный орган, например, легкие.
Возбудители гриппа относятся к семейству ортомиксовирусов (Orthomyxo-viridae) и являются пневмотропными вирусами. Схематическое строение вируса гриппа представлено на рис. 2.1. Геном вируса складывается из 8-ми фрагментов однонитчатой РНК, которые кодируют
10 вирусных белков. Фрагменты РНК имеют общую белковую оболочку, соединяющую их, образуя антигенно-стабильный рибонуклеопротеид (S-антиген), который определяет принадлежность вируса к серотипу А, В или С. Снаружи вирус покрыт двойным липидным слоем с внутренней стороны которого находится слой мембранного белка.
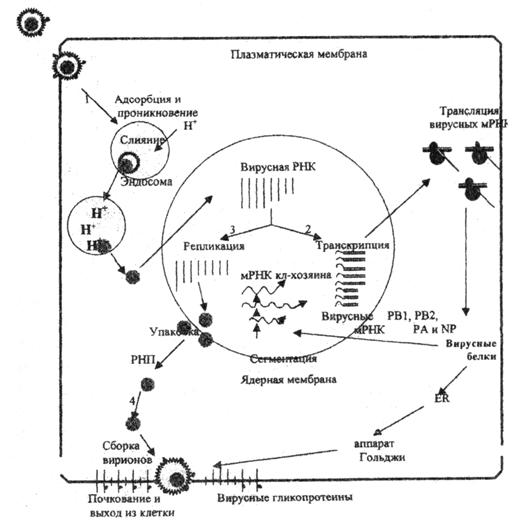
Рис. 1. Репликативный цикл вируса гриппа (В.И.Покровский, О.И.Киселев, 2002)
Представлены 4 основные фазы репликативного цикла вируса гриппа: 1 – адсорбция и проникновение вируса в клетку хозяина; 2 – транскрипция вирусной РНК и трансляция вирусных белков; 3 – репликация вирусной РНК и 4 – самосборка вирионов и последующий выход их из клетки.
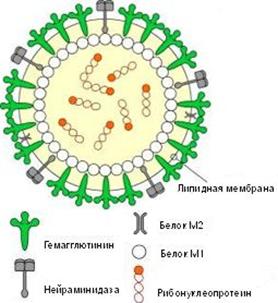
Рис. 2. Схематическое строение вируса гриппа
Гемаглютинин и нейраминидаза являются факторами агрессии вируса гриппа. Интенсивность интоксикации при гриппе определяется свойствами гемаглютинина, а нейраминидаза обладает выраженным иммунодепресивным действием. Оба поверхностных антигена характеризуются выраженной способностью к изменчивости, в результате чего появляются новые антигенные варианты вируса. Гемаглютинины 1, 2, 3 типов и нейраминидазы 1, 2 типов содержат вирусы, которые поражают человека. Другие антигены характерные для вирусов гриппа животных (свиней, собак, лошадей, многих видов птиц и др.).
Вирусы гриппа А. Эти возбудители более вирулентны и контагиозны, чем вирусы гриппа В и С. Это обусловлено тем, что вирус А содержит 2 типа нейраминидазы (N1, N2) и 4 –гемаглютинина (H0, H1, H2, H3); вирус В – 1 и 1 соответственно, поэтому он имеет лишь антигенные варианты внутри одного серотипа; вирус С содержит только гемаглютинин и не содержитнейраминидазу, но он имеет рецептор, который разрушает энзим. Вирусу С не свойственная изменчивость.
Одновременно циркулируют и имеют эпидемическое распространение 2 подтипа вируса гриппа А H3N2 и H1N1. На сегодняшний день во всем мире доминирует вариант А/Сидней/05/97, А/Берн/07/95 и А/Пекин/262/95. Характерной чертой современных вирусов гриппа А (H3N2) является изменение ряда их биологических свойств: тяжело размножаются в куриных эмбрионах, не агглютинируют эритроциты кур, взаимодействуют только с эритроцитами человека или морской свинки, имеют низкую иммуногенную активность.
Вирусы гриппа В выделяются только от людей. По вирулентности и эпидемиологической значимости они уступают вирусам гриппа А. По антигенным свойствам гемаглютинин инейраминидазу вируса В можно разделить на 5 подтипов. Их изменчивость носит более постепенный и медленный характер, что объясняет эпидемиологические особенности вируса гриппа В (эпидемии возникали 1 раз в 3-4 года до 1988 г.). В мире сейчас циркулируют 2 варианта вируса гриппа В: В/Пекин/184/93 (Европа, Америка, Африка, Австралия) и В/Виктория/2/87 (Юго-Восточная Азия). Характерной чертой современных вирусов гриппа А и В является их высокая чувствительность к ингибиторам гемаглютинирующей и инфекционной активности, которые содержатся в сыворотке крови нормальных животных.
Вирусы гриппа С . Вирусы гриппа С, в отличие от вирусов А и В, не вызывают эпидемий, но они могут быть причиной вспышек в организованных детских коллективах. Подъемы заболеваемости гриппом С нередко предшествуют или сопровождают эпидемии гриппа А и В. У детей раннего возраста и более старших возрастных группах он приводит к заболеваниям в легкой ибессимптомной форме.
Для вирусов гриппа С характерна значительно большая стабильность антигенных и биологических свойств. По ряду биологических характеристик эти возбудители отличаются от других представителей семейства ортомиксовирусов. Им присуща низкая репродуктивная активность в разных клеточных системах и наличие других, чем у вирусов гриппа А и В, рецепторов на поверхности эритроцитов. Рецептор-деструктирующая активность связана не с нейраминидазой, как у вирусов гриппа А и В, а с ферментом нейраминат-0-ацетилестеразою.
Сложность этиологии усиливают факты выделения от людей вирусов, которые не свойственные человеку. Гены вируса гриппа (H5N1), выделенные в 1997 году от человека в Гонконге, оказались подобными генам птичьего вируса, который преодолел межвидовой барьер. Оказалось, что человеческие и птичьи вирусы гриппа были реасортантами (рекомбинантные штаммы), которые получили внутренние гены РВ1 и РВ2 от перепелиного вируса гриппа А (H9N2) – штаммы А/перепел/Гонконг/61/97.
Вирусы гриппа малоустойчивы во внешней среде, в течение нескольких часов при комнатной температуре разрушаются. Под действием дезинфицирующих растворов (спирта, формалина, сулемы, кислот, щелочей) они быстро погибают. Нагревание до 50-60°С инактивирует вирусы в течение нескольких минут, в замороженном состоянии при температуре -70° С они сохраняются годами, не теряя инфекционных свойств, быстро погибают под воздействием ультрафиолетового излучения. Оптимум размножения вируса происходит при +37°С в слабощелочной среде. Как и другие вирусы, возбудитель гриппа отнюдь не чувствительный к антибиотикам и сульфаниламидам.
Вирус гриппа не растет на обычных питательных средах. Его можно выделить из материала, который получен от больного в первые дни болезни (смывы из носоглотки, мокроты), путем заражения культур или клеток куриных эмбрионов.
Традиционно считалось, что человеческий организм освобождается от возбудителя в ближайшие дни после выздоровления. Однако постепенно накапливались косвенные свидетельства, которые опровергают эту точку зрения. И в 1985 г. были получены прямые доказательства формирования длительной персистенции вируса гриппа. У одного человека удалось многократно выделять вирус гриппа в течение 9-ти месяцев и дольше. Возможность пожизненной персистенции вируса в организме человека продолжает изучаться.
1. Грипп и другие респираторные вирусные инфекции / под ред. О.И. Киселева, И.Г. Мариничева, А.А. Сомининой. – СПб. – 2003.
2. Дриневский В.П., Осидак Л.В., Цыбалова Л.М. Острые респираторные инфекции у детей и подростков // Практическое руководство под редакцией О.И. Киселева. – Санкт-Петербург. – 2003.
3. Железникова Г.Ф., Иванова В.В., Монахова Н.Е. Варианты иммунопатогенеза острых инфекций у детей. – Санкт-Петербург. – 2007. – 254 с.
4. Ершов Ф.И. Грипп и другие ОРВИ // Антивирусные препараты. Справочник. – М., 2006. –С. 226-247.
5. Ершов Ф.И., Романцов М.Г. Антивирусные средства в педиатрии. – М., 2005. – С.159-175.
6. Ершов Ф.И., Киселев О.И. Интерфероны и их индукторы (от молекул до лекарств). – М., 2005. – С. 287-292.
7. Иванова В.В. Острые респираторно-вирусные заболевания // Инфекционные болезни у детей. – М., 2002.
8. Онищенко Г.Г., Киселев О.И., Соминина А.А. Усиление надзора и контроля за гриппом как важнейший элемент подготовки к сезонным эпидемиям и очередной пандемии. – М., – 2004. – С.5-9.
9. Об утверждении стандарта медицинской помощи больным гриппом, вызванным идентифицированным вирусом гриппа (грипп птиц) // Приказ Минздравсоцразвития №460 от 07.06.2006 г.
10. Романцов М.Г., Ершов Ф.И. Часто болеющие дети: Современная фармакотерапия. – М., 2006. – 192 с.
11. Стандартизированные принципы диагностики, лечения и экстренной профилактики гриппа и других острых респираторных инфекций у детей / под ред. О.И. Киселева. – СПб. – 2004. – С. 82-95.
12. Лекарственные средства в фармакотерапии патологии клетки / под редакцией Т.Г. Кожока.-М., 2007.
Читайте также:
