
Патогенез вирусных болезней на клеточном уровне на уровне организма
Вирусы являются внутриклеточными инфекционными агентами. Весь репликативный цикл вируса осуществляется с исполь зованием метаболических и генетических ресурсов клеток. Поэтому патогенез вирусных инфекций, в первую очередь, следует рассматривать на молекулярном и клеточном уровнях (В.И. Покровский, О.И. Киселев, 2002). Вместе с тем, инфекционный процесс, вызванный вирусами, развивается в пределах того или иного органа или ткани, так как большинство вирусов обладают достаточно высокой органной или тканевой тропностью. Поэтому характер развития внутритканевых процес сов при вирусных инфекциях, с одной стороны, определяется, как правило, цитопатическим действием вируса на клетки данной ткани и органа, а с другой стороны, реакцией внутритканевых и органных систем защиты от вирусной инфекции. Последние не редко носят деструктивный характер и усугубляют течение забо левания и его последствия. Поэтому понимание патогенеза вирусных инфекций представляется исключительно важным для создания рациональных схем лечения, основанных на гармоничном сочетании симптоматического лечения и противовирусной химиотерапии.
Стадии репликативного цикла вируса гриппа представлены на рис. 1. Такой же репликативный цикл свойствен большинству РНК- и ДНК-содержащих вирусов, кроме тех, кото рые обладают особыми механизмами хронизации процесса, латенции и способностью к интеграции в клеточный геном (например, ВИЧ, гепатит С, вирус папилломы и.т.). Ключевыми стадиями репликативного цикла вируса являются проникновение в клетки, декапсидация, активация процессов транскрипции и трансляции, собственно репликация вирусного генома и созревание зрелых вирусных частиц с выходом нового потомства инфекционного вируса.
Проникновение вируса в клетки - одна из ключевых стадий патогенеза инфекционного процесса. Для большинства вирусов степень цитопатического действия вируса прямо зависит от множественности заражения. В естественных условиях инфицирова ние клеток вирусами не происходит обычно с высокой множе ственностью. Исключение составляют респираторные вирусы, которые передаются воздушно-капельным путем с высокой инфицирующей дозой. ВИЧ и гепатиты, наоборот, эффективно передаются с кровью в самых незначительных трудно определяе мых титрах. Такие вирусы имеют особые свойства, определяющие их контагиозность.
Для проникновения вируса в клетку, необходимо: высокое сродство к вирусоспецифическому рецептору; множественность рецепторов; эффективность слияния вируса с клетками при инфицировании; способность к образованию синцития, к образованию гигантских синпластов, состоящих из множества слившихся клеток, в которых вирусный нуклеопротеид без эндоцитоза беспрепятственно переходит от одной зараженной клетки к множеству других через цитоплазму.
Такими свойствами из респираторных вирусов обладает респираторно-синцитиальный вирус, а среди ретровирусов – вирус иммунодефицита человека.
Стадии жизненного цикла вируса гриппа и соответствующие ингибиторы репродукции в известной степени могут быть экстраполированы на многие другие вирусы, за исключением тех, жизненный цикл которых включает интеграцию в клеточный геном. В процессе репликации вирусов обычно происходит селективная блокада трансляции клеточных мРНК и активация экспресси клеточных генов, кодирующих провоспапительные лимфокины. Многократные раунды репликации вирусного генома приводят к истощению энер гетических ресурсов клеток и пула предшественников азотистых оснований. Повышенное содержание провоспалительных лимфокинов приводит к развитию воспалительной реакции и локально му генерированию свободных радикалов, оказывающих, в свою очередь, повреждающее действие на клетки и ткани через перекисное окисление липидов клеточных мембран, инактивацию жизненно важных ферментов. Поэтому одним из важнейших факторов патогенности вирусов является их репликативный потенциал: чем активнее происходит репликация, тем сильнее цитопатическое действие вирусов на клетки и пораженную ткань (орган).
Значительный вклад в цитопатическое действие вирусов вносит репродукция вирусов в митохондриях, что сопровождается нарушением функций этих органоидов, падением продукции АТФ и индукцией опосредованного митохондриями пути апоптоза.
Впервые возможность репродукции вирусов в митохондриях была доказана выдающимся отечественным вирусологом В.М. Ждановым. Узкий тропизм вирусов, в известной степени, спасителен для организма, так как инфекционный процесс ограничен одним органом.
Это обстоятельство позволяет, проводит рациональную симптоматическую терапию, направленную на защиту данной ткани или органа и ориентировать противовирусные средства в данный орган, например, легкие.
Возбудители гриппа относятся к семейству ортомиксовирусов (Orthomyxo-viridae) и являются пневмотропными вирусами. Схематическое строение вируса гриппа представлено на рис. 2.1. Геном вируса складывается из 8-ми фрагментов однонитчатой РНК, которые кодируют
10 вирусных белков. Фрагменты РНК имеют общую белковую оболочку, соединяющую их, образуя антигенно-стабильный рибонуклеопротеид (S-антиген), который определяет принадлежность вируса к серотипу А, В или С. Снаружи вирус покрыт двойным липидным слоем с внутренней стороны которого находится слой мембранного белка.
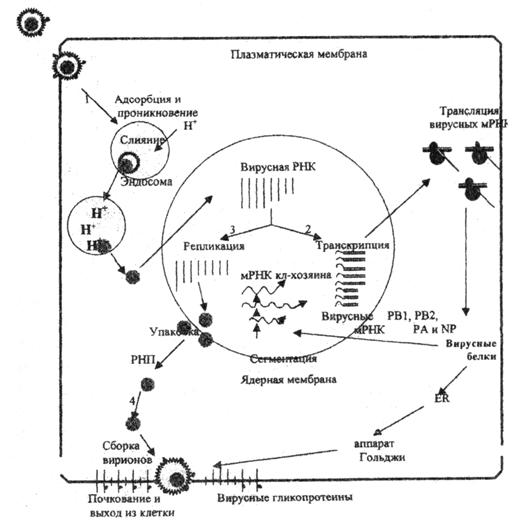
Рис. 1. Репликативный цикл вируса гриппа (В.И.Покровский, О.И.Киселев, 2002)
Представлены 4 основные фазы репликативного цикла вируса гриппа: 1 – адсорбция и проникновение вируса в клетку хозяина; 2 – транскрипция вирусной РНК и трансляция вирусных белков; 3 – репликация вирусной РНК и 4 – самосборка вирионов и последующий выход их из клетки.
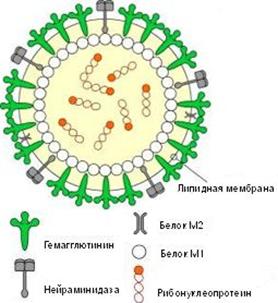
Рис. 2. Схематическое строение вируса гриппа
Гемаглютинин и нейраминидаза являются факторами агрессии вируса гриппа. Интенсивность интоксикации при гриппе определяется свойствами гемаглютинина, а нейраминидаза обладает выраженным иммунодепресивным действием. Оба поверхностных антигена характеризуются выраженной способностью к изменчивости, в результате чего появляются новые антигенные варианты вируса. Гемаглютинины 1, 2, 3 типов и нейраминидазы 1, 2 типов содержат вирусы, которые поражают человека. Другие антигены характерные для вирусов гриппа животных (свиней, собак, лошадей, многих видов птиц и др.).
Вирусы гриппа А. Эти возбудители более вирулентны и контагиозны, чем вирусы гриппа В и С. Это обусловлено тем, что вирус А содержит 2 типа нейраминидазы (N1, N2) и 4 –гемаглютинина (H0, H1, H2, H3); вирус В – 1 и 1 соответственно, поэтому он имеет лишь антигенные варианты внутри одного серотипа; вирус С содержит только гемаглютинин и не содержитнейраминидазу, но он имеет рецептор, который разрушает энзим. Вирусу С не свойственная изменчивость.
Одновременно циркулируют и имеют эпидемическое распространение 2 подтипа вируса гриппа А H3N2 и H1N1. На сегодняшний день во всем мире доминирует вариант А/Сидней/05/97, А/Берн/07/95 и А/Пекин/262/95. Характерной чертой современных вирусов гриппа А (H3N2) является изменение ряда их биологических свойств: тяжело размножаются в куриных эмбрионах, не агглютинируют эритроциты кур, взаимодействуют только с эритроцитами человека или морской свинки, имеют низкую иммуногенную активность.
Вирусы гриппа В выделяются только от людей. По вирулентности и эпидемиологической значимости они уступают вирусам гриппа А. По антигенным свойствам гемаглютинин инейраминидазу вируса В можно разделить на 5 подтипов. Их изменчивость носит более постепенный и медленный характер, что объясняет эпидемиологические особенности вируса гриппа В (эпидемии возникали 1 раз в 3-4 года до 1988 г.). В мире сейчас циркулируют 2 варианта вируса гриппа В: В/Пекин/184/93 (Европа, Америка, Африка, Австралия) и В/Виктория/2/87 (Юго-Восточная Азия). Характерной чертой современных вирусов гриппа А и В является их высокая чувствительность к ингибиторам гемаглютинирующей и инфекционной активности, которые содержатся в сыворотке крови нормальных животных.
Вирусы гриппа С . Вирусы гриппа С, в отличие от вирусов А и В, не вызывают эпидемий, но они могут быть причиной вспышек в организованных детских коллективах. Подъемы заболеваемости гриппом С нередко предшествуют или сопровождают эпидемии гриппа А и В. У детей раннего возраста и более старших возрастных группах он приводит к заболеваниям в легкой ибессимптомной форме.
Для вирусов гриппа С характерна значительно большая стабильность антигенных и биологических свойств. По ряду биологических характеристик эти возбудители отличаются от других представителей семейства ортомиксовирусов. Им присуща низкая репродуктивная активность в разных клеточных системах и наличие других, чем у вирусов гриппа А и В, рецепторов на поверхности эритроцитов. Рецептор-деструктирующая активность связана не с нейраминидазой, как у вирусов гриппа А и В, а с ферментом нейраминат-0-ацетилестеразою.
Сложность этиологии усиливают факты выделения от людей вирусов, которые не свойственные человеку. Гены вируса гриппа (H5N1), выделенные в 1997 году от человека в Гонконге, оказались подобными генам птичьего вируса, который преодолел межвидовой барьер. Оказалось, что человеческие и птичьи вирусы гриппа были реасортантами (рекомбинантные штаммы), которые получили внутренние гены РВ1 и РВ2 от перепелиного вируса гриппа А (H9N2) – штаммы А/перепел/Гонконг/61/97.
Вирусы гриппа малоустойчивы во внешней среде, в течение нескольких часов при комнатной температуре разрушаются. Под действием дезинфицирующих растворов (спирта, формалина, сулемы, кислот, щелочей) они быстро погибают. Нагревание до 50-60°С инактивирует вирусы в течение нескольких минут, в замороженном состоянии при температуре -70° С они сохраняются годами, не теряя инфекционных свойств, быстро погибают под воздействием ультрафиолетового излучения. Оптимум размножения вируса происходит при +37°С в слабощелочной среде. Как и другие вирусы, возбудитель гриппа отнюдь не чувствительный к антибиотикам и сульфаниламидам.
Вирус гриппа не растет на обычных питательных средах. Его можно выделить из материала, который получен от больного в первые дни болезни (смывы из носоглотки, мокроты), путем заражения культур или клеток куриных эмбрионов.
Традиционно считалось, что человеческий организм освобождается от возбудителя в ближайшие дни после выздоровления. Однако постепенно накапливались косвенные свидетельства, которые опровергают эту точку зрения. И в 1985 г. были получены прямые доказательства формирования длительной персистенции вируса гриппа. У одного человека удалось многократно выделять вирус гриппа в течение 9-ти месяцев и дольше. Возможность пожизненной персистенции вируса в организме человека продолжает изучаться.
1. Грипп и другие респираторные вирусные инфекции / под ред. О.И. Киселева, И.Г. Мариничева, А.А. Сомининой. – СПб. – 2003.
2. Дриневский В.П., Осидак Л.В., Цыбалова Л.М. Острые респираторные инфекции у детей и подростков // Практическое руководство под редакцией О.И. Киселева. – Санкт-Петербург. – 2003.
3. Железникова Г.Ф., Иванова В.В., Монахова Н.Е. Варианты иммунопатогенеза острых инфекций у детей. – Санкт-Петербург. – 2007. – 254 с.
4. Ершов Ф.И. Грипп и другие ОРВИ // Антивирусные препараты. Справочник. – М., 2006. –С. 226-247.
5. Ершов Ф.И., Романцов М.Г. Антивирусные средства в педиатрии. – М., 2005. – С.159-175.
6. Ершов Ф.И., Киселев О.И. Интерфероны и их индукторы (от молекул до лекарств). – М., 2005. – С. 287-292.
7. Иванова В.В. Острые респираторно-вирусные заболевания // Инфекционные болезни у детей. – М., 2002.
8. Онищенко Г.Г., Киселев О.И., Соминина А.А. Усиление надзора и контроля за гриппом как важнейший элемент подготовки к сезонным эпидемиям и очередной пандемии. – М., – 2004. – С.5-9.
9. Об утверждении стандарта медицинской помощи больным гриппом, вызванным идентифицированным вирусом гриппа (грипп птиц) // Приказ Минздравсоцразвития №460 от 07.06.2006 г.
10. Романцов М.Г., Ершов Ф.И. Часто болеющие дети: Современная фармакотерапия. – М., 2006. – 192 с.
11. Стандартизированные принципы диагностики, лечения и экстренной профилактики гриппа и других острых респираторных инфекций у детей / под ред. О.И. Киселева. – СПб. – 2004. – С. 82-95.
12. Лекарственные средства в фармакотерапии патологии клетки / под редакцией Т.Г. Кожока.-М., 2007.
Патогенез на клеточном уровне.
Патогенез любой вирусной инфекции завершается общим поражением организма, а начинается он на клеточном уровне, что обусловлено облигатным внутриклеточным паразитизмом вирусов.
Цитопатический эффект вируса – специфическая морфологическая деструкция и функциональная патология клеток, культивируемых вне организма, вызванная заражением их цитопатогенными вирусами. Различают три типа ЦПД:
цитолитический – характеризуется лизисом клеток. Дегенерирующие клетки отслаиваются от стекла, через 20-26 ч культура представляет собой сеть синцитиальных элементов, а через 48 ч на стекле сохраняются единичные веретенообразные и отростчатые клетки;
трансформирующий – не вызывает гибели клеток, а дает им способность к неограниченному размножению;
индуктивный – характеризуется способностью инфицированных клеток продуцировать интерферон.
ЦПД зависит от вида вируса и чувствительности к нему культуры клеток. Образование симпластов идет по двум основным механизмам:
разрушение клеточных стенок и слияние близлежащих клеток (с участием ферментов внешних оболочек вирусов, например нейраминидазы);
амитоз без деления цитоплазмы.
ЦПД преимущественно связано с повреждением клеточных оболочек энзимами вирусов. Нейтрализация ЦПД специфической сывороткой – важный диагностический метод, позволяющий идентифицировать вирус, индуцирующий ЦПД.
При ряде вирусных инфекций в клетках появляются характерные для данного заболевания клеточные включения. Их обнаружение имеет диагностическое значение (бешенство, оспа, аденовирусы и др.). Включения могут локализоваться в цитоплазме или ядре, а по своей природе быть либо скоплениями вирионов, либо продуктом реакции клетки на вирусную инфекцию.
Патогенез на уровне организма.
Пути проникновения вирусов в организм разнообразны. Вирусы могут попадать в организм через носоглотку, ЖКТ, при прямом кожном контакте, передаваться через эктопаразитов.
Диссеминация вирусов в организме.
После проникновения вируса в организм он может размножаться вначале на месте проникновения или в определенных органах и тканях и далее разноситься кровью по всему организму.
В основном диссеминация вирусов осуществляется лимфо- и гематогенно. Нейротропные вирусы диссеминируют по нервным стволам.
Выделение вируса из организма.
При заболеваниях, вызванных пантропными вирусами, выделение идет с фекалиями, мочой, экссудатом, молоком, слюной. При гриппе, парагриппе, ИРТ КРС вирус может выделяться через носоглотку и с фекалиями. При аденоинфекции вирус выделяется с носоглоточным секретом, фекалиями, мочой; при энтеровирусных заболеваниях - гл. образом с фекалиями; при болезнях, характеризующихся кожными поражениями – обычно из этих поражений; при бешенстве – в основном со слюной. Возможно выделение вируса со спермой (ящур, лейкоз, ИРТ, ВД).
Предпочтительная локализация вируса в инфицированном организме.
Тропизм (аффинитет) вируса определяется его поверхностными структурами. Избирательная локализация вирусов происходит в тех клетках и тканях, где может реализоваться генетическая информация вируса, хотя все вирусы широко распространяются в организме.
По тропизму вирусы разделяют на следующие группы:
Однако классификация вирусов на основе тропизма невозможна, т.к. это сродство нестрогое и один и тот же вирус может вызывать несколько клинических форм болезни.
Влияние вирусов на кровь.
В период максимальной инфекционности крови проявляется феномен эритроцитотропизма – в эритроцитах обнаруживаются ДНК-содержащие включения. Помимо этого, при чуме свиней происходит торможение продукции эритроцитов и гранулоцитов. Задолго до появления клинических признаков болезни возникает деструкция кроветворных клеток, нарушение их синтеза и созревания, аплазия.
Лейкоциты могут адсорбировать на себе вирусы, из-за чего снижается их фагоцитирующая активность. При чуме и ВД постоянным признаком является лейкопения. Снижается интерферосинтезирующая активность лейкоцитов.
Степень патогенности вирусов зависит от особенностей штамма и метода его поддержания, проявление ее – от метода введения вируса в организм. Различные штаммы одного и того же вируса могут различаться по своей вирулентности (у болезни Ньюкасла – вело-, мезо- и лентогенные штаммы).
Факторы, влияющие на чувствительность животных к вирусам.
Существует пять таких факторов:
Из гормонов наибольшее значение имеет кортизон, а т.ж. тестостерон. Избыток кортизона в основном усиливает действие вирусов.
Роль возраста в патогенезе вирусных инфекций в значительной степени связана с влиянием гормонов (известны факты большей восприимчивости и, наоборот, большей резистентности молодняка к вирусным инфекциям).
Подавляющее большинство врожденных пороков развития обусловлено вирусами, вызывающими различные болезни. Иммунизация беременных самок может привести к тому же результату (тератогенному эффекту или мертворождению). Многие вирусы могут передаваться трансплацентарно.
Под влиянием ионизирующего излучения изменяется чувствительность организма к патогенному действию некоторых вирусов. Рентгеновское облучение ухудшает течение многих вирусных инфекций. Под его влиянием повышается проницаемость сосудов и тканей, угнетается защитная роль барьерных систем, снижается бактерицидность тканей и крови. В облученных клетках резко нарушается течение основных метаболических процессов, особенно матричная активность ДНК.
Влияние температуры на чувствительность организма к вирусной инфекции проявляется неоднозначно. В развитии первичной вирусной инфекции играют роль в основном три фактора: повышение температуры, снижение рН в некоторых тканях и продукция эндогенного интерферона. Гипертермия организма оказывает губительное воздействие на низкопатогенные штаммы вирусов, влияя в первую очередь на размножение.
Зачастую бессимптомные формы вирусной инфекции вызывают живые вакцинные штаммы (против чумы КРС, чумы плотоядных и т.д.). судить о наличии возбудителя в организме животного при латентной инфекции можно по парным сывороткам.
Некогда тяжелые в пошлом болезни, оставлявшие после себя прочный иммунитет, в настоящее время снизили свою тяжесть и часто вызывают бессимптомные затяжные инфекции. Часто бессимптомная инфекция связана с персистенцией.
В патогенезе РВИ выделяют пять основных фаз:
Репродукция вируса в клетках ВДП в результате заражения или активации латентной инфекции, вызванной др. вирусами.
Вирусная и токсико-аллергическая реакции со стороны различных систем и внутренних органов; возможна вирусемия.
Поражение дыхательных путей с преимущественной локализацией процесса в различных отделах дыхательного тракта.
Бактериальные осложнения со стороны дыхательных путей (наиболее длительная и тяжелая фаза).
Обратное развитие патологического процесса с возможным формированием латентных или хронических форм.
Патогенез вакцинальной инфекции.
При вакцинации живыми противовирусными вакцинами возможны поствакцинальные осложнения, наиболее опасными из которых являются неврологические – в виде энцефалитов и менингоэнцефалитов.
Методом борьбы с поствакцинальной инфекцией служит разработка субъединичных вакцин.
Хронические вирусные инфекции.
Существует гипотеза о том, что персистенция вируса обусловлена блоком созревания вируса, в результате чего в клетках накапливаются нуклеокапсиды. В соответствии с этой гипотезой принципиальным отличием хронически инфицированных клеток от клеток с острой инфекцией является отсутствие сборки вирионов. Устранение такой блокировки должно приводить к переходу хронической инфекции в острую.
Установлено различие в свойствах нуклеокапсидов при персистенциях и острых инфекциях. Исходя из этого пытаются объяснить патогенез некоторых хронических инфекций. Вирус проникает в организм при острой инфекции или вакцинации, но острый процесс прерывается антителами и вирус элиминируется. Однако в результате вирусемии вирус контактирует с клетками различных систем, в том числе и с малочувствительными, проникая в них. В результате абортивного цикла репродукции сборки вирионов в клетках не происходит, но идет накопление субвирусных компонентов и нуклеокапсидов. Из-за наличия в клетке вирусного гликопротеида, вызывающего слияние, происходит образование симпластов с проникновением нуклеокапсида в новые клетки. Этим обуславливается медленное течение и неизбежный трагический исход болезни.
Аутоимунные вирусные болезни.
При протекании болезней этой группы иммунная система распознает как чужие некоторые собственные клеточные и тканевые антигены, в результате чего иммунной атаке подвергаются неизменные клетки и ткани организма хозяина.
Наиболее подробно изучено действие системы вирус – антитело – комплемент.
В условиях промышленного животноводства проблема вирусобактериальных инфекций приобретает особую актуальность.
Иммунодефицит вирусной природы.
Состояние иммунной недостаточности может быть обусловлено рядом вирусов и генетической предрасположенностью. Приобретенные иммунодефициты могут являться результатом воздействия вируса, иногда скрытого. При этом может снижаться эффективность противовирусных вакцин. Некоторые вирусы вызывают лимфопению, а острая форма болезни Марека полностью прекращает лимфопоэз, следствием чего является депрессия гуморального ответа на гетерологичные антигены.
Особенно чувствительна к вирусам система мононуклеарных фагоцитов, часто являющаяся мишенью для прямого действия вирусов.
Ветеринарная вирусология. Сюрин В.Н., Белоусова Р.В., Фомина Н.В. – М.: Агропромиздат, 1991. – стр. 222 – 249.








ПАТОГЕНЕЗ ВИРУСНЫХ ИНФЕКЦИЙ
1. Автономная: Продуктивная (острая, хроническая, цитологическая, не цитологическая), Абортивная (острая, хроническая);
2. Интеграционная: Интеграция полного генома (неопластическая трансформация), Интеграционные части (отсутствие трансформации) и течения, а также тяжелым восприятием организма на заражение вирусной болезнью, а иногда и бессимптомным.
Патогенез любой вирусной инфекции завершается общим поражением организма, а начинается он на клеточном уровне, что обусловлено облигатным внутриклеточным паразитизмом вирусов. ЦПД зависит от вида вируса и чувствительности к нему культуры клеток. Образование симпластов, связано с повреждением клеточных оболочек энзимами вирусов.
При ряде вирусных инфекций в клетках появляются характерные для данного заболевания клеточные включения. Их обнаружение имеет диагностическое значение (бешенство, оспа, аденовирусы и др.). Включения могут локализоваться в цитоплазме или ядре, а по своей природе быть либо скоплениями вирионов, либо продуктом реакции клетки на вирусную инфекцию.
Пути проникновения вирусов в организм разнообразны. Вирусы могут попадать в организм через носоглотку, ЖКТ, при прямом кожном контакте, передаваться через эктопаразитов. При заболеваниях, вызванных пантропными вирусами, выделение идет с фекалиями, мочой, экссудатом, молоком, слюной. При гриппе, парагриппе, ИРТ КРС вирус может выделяться через носоглотку и с фекалиями. При аденоинфекции вирус выделяется с носоглоточным секретом, фекалиями, мочой; при энтеровирусных заболеваниях - гл. образом с фекалиями; при болезнях, характеризующихся кожными поражениями – обычно из этих поражений; при бешенстве – в основном со слюной. Возможно выделение вируса со спермой (ящур, лейкоз, инфекционный ринотрахеит, вирусная диарея).
Динамика инфекционной болезни:
1-й инкубационный ,или скрытый период (от момента проникновения возбудителя в организм или ткани, до появления первых клинических признаков).
2-й предклинический период (продолжительность от момента проявления признаков до их развития (1-2 дня). Появление общих не специфических признаков болезни).
3-й период полного развития болезни (активное выделение вируса).
4-й период угасания (клиническое выздоровление, или смерть).
Возможные исходы вирусных болезней:
1) Цитопагенное действие вируса; 2) трансформация клеток; 3) развитие латентной формы инфекции.
Лабораторный диагноз на патогенную инфекционную болезнь устанавливают на основании серологических (реакция диффузионной преципитации), случаях – ставят биологическую пробу.
Понимание патогенеза вирусных инфекций представляется исключительно важным для создания рациональных схем лечения, основанных на гармоничном сочетании симптоматического лечения и противовирусной химиотерапии. Смешанные инфекции представляют собой опасный путь развития процесса.
Библиографический список:
Ветеринарная вирусология: учебник для вузов / Р. В. Белоусова, Э. А. Преображенская, И. В. Третьякова; под ред. Р. В. Белоусовой. - М. : КолосС , 2007. - 424 с.
Ветеринарная вирусология: [учебник для вузов] / Р. Г. Госманов, Н. М. Колычев. - 2-е изд., перераб. и доп. - М. : КолосС, 2006. - 304 с.,
Патологическая физиология / Адо А.Д. и др. М., 2000.
Давыдовский, И.В. Общая патология человека, М., 2003.-290с.
Еще на заре существования человечества людей чрезвычайно интересовали механизмы возникновения болезней.
Причиной заболевания сначала считали исключительно внешние болезнетворные факторы: простуду, голод и даже сверхъестественные силы. Открытия более поздних лет позволили точнее формулировать взгляды на внешние причины заболевания, отнеся их главным образом за счет воздействия болезнетворных организмов. Но и в те времена существовали предпосылки против столь одностороннего понимания причин даже инфекционных болезней. И только с развитием молекулярной биологии поиск причин заболевания стал включать в себя не только изучение вредных факторов внешней среды, но и сам организм.
Под инфекцией в настоящее время понимают состояние зараженности организма с антагонистическими взаимоотношениями между патогенным возбудителем и хозяином. Оно проявляется в виде болезни или заразоносительства.
Вирусные болезни передаются от больных животных здоровым различными путями. Например, аэрогенно передаются вирусы инфекционного бронхита кур, инфекционного ринотрахеита крупного рогатого скота, оспы. Алиментарно, т. е. с кормом, с водой, передаются вирусы ящура, вирусной диареи крупного рогатого скота, гриппа птиц. Контактно, т. е. прямым соприкосновением, или непрямым контактом — через инфицированные предметы ухода, передается вирус оспы. Около 40 % вирусов передается трансмиссивно, т. е. насекомыми. Например, вирус инфекционной анемии лошадей переносится комарами, клещевого энцефалита — клещами, с укусом передается вирус бешенства. И наконец, трансовариально передается вирус инфекционного бронхита кур, трансплацентарно — вирус инфекционного гастроэнтерита свиней, вирусной диареи крупного рогатого скота.
Началом инфекционного процесса служит проникновение вируса в организм. Место проникновения зависит от способа передачи вируса и локализации чувствительных к данному вирусу клеток.
Большая часть вирусов проникает в организм хозяина через барьеры слизистых оболочек дыхательных путей и пищеварительного тракта. Поверхность слизистых оболочек защищена от вирусов комплексом факторов: неспецифическими ингибиторами, протеолитическими ферментами, слизью, солями желчных кислот, лизоцимом, Е-киллерами. Заражение произойдет, если вирус окажется резистентен к названным факторам.
Вирусы оспы способны проникать через неповрежденную кожу, а вирус классической чумы свиней, инфекционной анемии лошадей — через поврежденную кожу с укусом членистоногими, вирус иммунодефицита человека — при переливании крови.
Нахождение вируса на месте внедрения называют первичной локализацией вируса, которая может быть в эпителии кожи или слизистых оболочек, в регионарных лимфатических узлах, в клетках тканей. Некоторые вирусы, например гриппа, в месте первичной локализации уже размножаются. Тогда в ответ на размножение вируса в организме синтезируется интерферон как фактор естественной видовой и неспецифической резистентности.
Следующим этапом развития вирусной болезни является первичная циркуляция вируса по организму (диссеминация), которая может осуществляться с током крови (виремия), лимфы или по нервным стволам. Вирусы бешенства, болезни Борна, алеутской болезни норок распространяются по нервам в восходящем (центростремительном) направлении. Такую миграцию вируса называют нейропробазией.
Распространившись по организму, вирус локализуется в чувствительных к нему клетках определенного типа. Чувствительной к вирусу является та клетка, которая имеет на поверхности рецепторы, комплементарные данному вирусу и необходимые для адсорбции на ней вируса; в цитоплазме чувствительных к вирусу клеток находятся ферменты для его депротеинизации. По тому, в каких клетках организма размножаются и локализуются те или иные вирусы, их условно разделяют на нейро-, дерма-, пневмо-, энтеро-, висцеро — и пантропные.
Размножение вируса в клетках ведет к изменению их обмена веществ, морфологии и функции. В результате появляются цитопатическое действие вируса и вирусные тельца-включения. Непосредственными причинами, ведущими к повреждению и даже гибели пораженных вирусом клеток, мот быть: механическое воздействие массы вирионов (вплоть до разрыва клетки); токсическое действие промежуточных и структурных белков; интеграция вирусного генома с клеточным; повреждение клеточных лизосом и митохондрий; паралич клеточного генома.
Одним из видов патологии клеток, зараженных вирусом, является их трансформация (в культуре клеток) или малигнизация (в организме). В результате действия вируса изменяются поверхностные свойства клеток, что ведет к их неограниченному делению, нарушению связей с окружающими клетками и распространению с током крови и лимфы по организму. В месте задержки такой клетки происходит ее активное проникновение между нормальными клетками и размножением. Возникает метастаз, а затем опухоль.
На основании ряда исследований сформулирована гипотеза, по которой клетки большинства позвоночных несут вертикально передаваемую информацию вируса, встроенную в ДНК клетки. Она и служит источником трансформации нормальной клетки в опухолевую. Фенотипическое проявление этого процесса может происходить или не происходить, что зависит от состояния организма и определенных внешних воздействий.
Способностью вызывать неограниченный рост клеток обладают как ДНК-содержащие вирусы (адено-, герпес-, паповавирусы), так и PH К-содержащие (ретровирусы). Таким вирусам дали название — онкогенные.
По мере нарастания числа пораженных вирусом клеток развиваются патологические изменения в функционировании органа, что ведет к появлению определенных клинических признаков заболевания и патологических изменений органов и тканей. Время от момента заражения до появления клинических симптомов называется инкубационным периодом.
Одновременно с развитием болезни включается клеточный противовирусный иммунитет — активизируются и клонируются Т — и В-лимфоциты. Это приводит к разрушению зараженных вирусом клеток и появлению гуморального иммунитета — противовирусных антител.
В период поражения вирусом органов и тканей ослабевает иммунитет организма, и тогда наступает обострение бактериальных инфекций. Именно наличие таких инфекций делает течение болезни тяжелым и нередко приводит к гибели.
В зависимости от локализации патологического процесса вирус выделяется из организма различными путями: с отделяемым слизистых оболочек носа и глаз, со слюной, с молоком, со спермой, с мочой, фекалиями, корочками кожных поражений. Это необходимо учитывать при проведении мероприятий по борьбе с тем или иным заболеванием.
Животные являются вирусоносителями в течение всей болезни и в разный период времени после переболевания (реконвалесцентное вирусоносительство). Вирусоносители периодически могут быть вирусовыделителями, что имеет большое значение для сохранения стационарного неблагополучия хозяйства.
В зависимости от вирулентности возбудителя и реактивности организма длительность нахождения вируса в организме различна. Короткое время вирус присутствует в организме при инапарантной и острой инфекции. Если вирус авирулентный, а животное малочувствительно к нему, то клинических признаков болезни не отмечают и вскоре происходит освобождение организма от вируса. Это инаппарантная инфекция.
Острая инфекция характеризуется ярким проявлением клинических признаков и заканчивается выздоровлением или гибелью. Выздоровление наступает вследствие действия комплекса факторов, иммунитета и интерференции между формами вируса — стандартными вирионами и дефектными интерферирующими частицами. Значительно реже наступает гибель животного. Причиной ее может быть повреждение жизненно важных органов (мозга, сердца), обширные повреждения нескольких органов, вызванных действием пантропных вирусов (чума собак) и развитием осложнений, вызванных бактериальной микрофлорой.
Длительное пребывание вируса в организме носит название персистенция (от лат. persistentia — постоянство). Персистенцией объясняется механизм латентной, хронической и медленной инфекций.
Латентная инфекция — это бессимптомная персистенция вируса; вирус может выделяться во внешнюю среду, а под действием стрессов возможно развитие острой инфекции (например, герпесвирусная инфекция).
Хроническая инфекция представляет собой персистенцию вируса, характеризующуюся чередованием периодов обострений и ремиссий, т. е. бессимптомных периодов, на протяжении недель или месяцев. Хроническая инфекция, как правило, заканчивается выздоровлением (например, аденовирусная инфекция или инфекционный ринотрахеит крупного рогатого скота).
Медленная инфекция имеет длительный инкубационный период — от нескольких недель до нескольких лет. Поражается обычно один орган, а развитие симптомов заболевания приводит к гибели. Такими заболеваниями являются, например, алеутская болезнь норок, инфекционная анемия лошадей, СПИД. Патогенез медленных инфекций до сих пор до конца не изучен и представляет собой различные механизмы патологии иммунной системы.
Если вы нашли ошибку, пожалуйста, выделите фрагмент текста и нажмите Ctrl+Enter.
Читайте также:
