
Интегративная вирусная инфекция это
Фазы взаимодействия вируса с клеткой:
1. Адсорбция вируса на клетке. В основе этого процесса лежит взаимодействие вирусных рецепторов со специфическими рецепторами клеток микроорганизма.
2. Проникновение вируса в клетку:
а) через слияние суперкапсида вируса с мембраной клетки. В результате этого слияния в цитоплазму клеток попадает только нуклеокапсид, а суперкапсидная оболочка остается на поверхности клетки;
б) путем эндоцитоза (пиноцитоза) с последующим освобождением вируса от суперкапсида в цитоплазме клеток.
3. Эклипс - фаза (фаза тумана). Происходит полная депротеинизация вируса, т.е. освобождение вирусной нуклеиновой кислоты от капсидной оболочки. В эту фазу вирус теряет свою морфологию и становится невидимым при электронной микроскопии.
4. Синтез (репликация) вирусных частиц:
Типы вирусной инфекции на уровне клетки:
1. Продуктивная.а) продуктивная острая - вирус внедряется в клетку, размножается, затем одномоментно выходит из клетки в больших количествах. При этом клетка сразу погибает.
б) продуктивная хроническая - вирус внедряется в клетку, размножается, затем постепенно выходят из клетки. При этом клетка длительно продуцирует вирусы, пока не будут истощены ее резервы. Затем клетка погибает.
а) для ДНК-содержаших вирусов: происходит интеграция ДНК вируса с ДНК клеткихозяина.
28.Интегративная вирусная инфекция. Онкогенные вирусы, классификация, вызываемые заболевания. Вирусо-генетическая теория онкогенеза ЛА.Зильбера.
В 50-х годах XX столетия Л.А.Зильбер сформулировал вирусо-генетическую теорию злокачественных опухолей, согласно которой вирусная ДНК интегрирует с ДНК клетки-хозяина, что приводит к трансформации нормальных клеток в опухолевые. Механизм интеграции для ДНК- и РНК-содержащих онкогенных вирусов представлен выше. Дальнейшие исследования показали, что в генетическом аппарате опухолевых вирусов есть особый ген (онкоген), который отвечает за синтез опухолевого белка, иначе Т-антигена (от лат. tumor - опухоль). Именно под действием Т-антигена в трансформированных клетках появляются следующие свойства:
а) способность клеток к непрерывному, неограниченному делению, т.к. между клетками нарушается контактное торможение;
б) способность клеток к делению в незрелом состоянии.
От момента интеграции вирусной ДНК с геномом клетки-хозяина до появления опухоли может пройти достаточно длительный период времени, в течение которого вирус не проявляет никакого трансформирующего действия на клетку. Но под влиянием химических канцерогенов, ионизирующего излучения, стрессовых
факторов, при старении организма происходит активация онкогена и возникает опухоль.
Классификация онкогенных вирусов:
I. ДНК-содержащие вирусы.
1. Вирусы группы оспы (поксвирусы) - вызывают доброкачественные опухоли у животных и человека. Например, вирус контагиозного моллюска вызывает доброкачественные опухоли кожи у человека
а) вирус простого герпеса II типа вызывает рак шейки матки;
б) вирус Эпштейна-Барр - возбудитель лимфомы Беркитта и назофарингеально го рака.
3. Паповавирусы (вирусы папиллом у животных и человека, вирусы полиомы у
грызунов и вакуолизирующий вирус SV 40 - вызывает саркому у хомяков и крыс).
4. Аденовирусы вызывают злокачественные опухоли у новорожденных хомяков.
5. Вирус гепатита В - возбудитель первичного рака печени.
П. РНК-содержашие онкогенные вирусы (ретровирусы) - вызывают саркомы, лейкозы, рак молочных желез у животных и человека.
Дата публикования: 2015-03-29 ; Прочитано: 1630 | Нарушение авторского права страницы
studopedia.org - Студопедия.Орг - 2014-2020 год. Студопедия не является автором материалов, которые размещены. Но предоставляет возможность бесплатного использования (0.002 с) . 
Вирусы - облигатные внутриклеточные паразиты, способные размножаться только в живой клетке. В отличие от прокариотических и эукариотических микроорганизмов вирусы не размножаются бинарным делением. Размножение вирусов происходит путём репродукции (англ, "reproduce" - воспроизводить, делать копию), то есть воспроизведение их нуклеиновых кислот и белков z последующей сборкой вирионов. Синтез нуклеиновых кислот и белков вируса происходит в разных частях клетки (ядре и цитоплазме). Такой способ репродукции получил название дизъюнктивного (разобщённого).
Процесс репродукции вирусов условно можно разделить на 2 фазы. Первая фаза включает 3 стадии:
- адсорбцию вируса на чувствительных клетках;
- проникновение вируса в клетку;
- депротеинизацию вируса.
Вторая фаза включает стадии реализации вирусного генома:
- транскрипцию,
- трансляцию,
- репликацию,
- сборку, созревание вирусных частиц,
- выход вируса из клетки.
Взаимодействие вируса с клеткой начинается с процесса адсорбции, т. е. с прикрепления вируса к поверхности клетки.
Адсорбция представляет собой специфическое связывание вирионного белка (антирецептора) с комплементарной структурой клеточной поверхности - клеточным рецептором. По химической природе рецепторы, на которых фиксируются вирусы, относятся к двум группам: мукопротеидным и липопротеидным. Вирусы гриппа, парагриппа, аденовирусы фиксируются на мукопротеидных рецепторах. Энтеровирусы, вирусы герпеса, арбовирусы адсорбируются на липопротеидных рецепторах клетки.
Адсорбция происходит лишь при наличии определённых электролитов, в частности ионов Са2+, которые нейтрализуют избыточные анионные заряды вируса и клеточной поверхности и уменьшают электростатическое отталкивание Адсорбция вирусов мало зависит от температуры Начальные процессы адсорбции носят неспецифический характер, являются результатом электростатического взаимодействия положительно и отрицательно заряженных структур на поверхности вируса и клетки, а затем наступает специфическое взаимодействие прикрепительного белка вириона со специфическими группировками на плазматической мембране клетки.
Простые вирусы человека и животных содержат прикрепительные белки в составе капсида. У сложно организованных вирусов прикрепительные белки входят в состав суперкапсида. Они могут иметь форму нитей (фибры у аденовирусов), либо шипов, грибоподобных структур у миксо-, ретро-, рабдо- и других вирусов. Вначале происходит единичная связь вириона с рецептором - такое прикрепление непрочное - адсорбция носит обратимый характер. Чтобы наступила необратимая адсорбция, должны появиться множественные связи между рецептором вируса и рецептором клетки, т. е. стабильное мультивалентное прикрепление. Количество специфических рецепторов на поверхности одной клетки составляет 104-105. Рецепторы для некоторых вирусов, например, для арбовирусов. содержатся на клетках как позвоночных, так и беспозвоночных, для других вирусов только на клетках одного или нескольких видов.
Проникновение вирусов человека и животных в клетку происходит двумя путями:
- виропексисом (пиноцитозом);
- слиянием вирусной суперкапсидной оболочки е клеточной мембраной.
Бактериофаги имеют свой механизм проникновения, так называемый шприцевой, когда в результате сокращения белкового отростка фага нуклеиновая кислота как бы впрыскивается в клетку.
Депротеинизация вируса освобождение генома вируса от вирусных защитных оболочек происходит либо с помощью вирусных ферментов, либо с помощью клеточных ферментов. Конечными продуктами депротеинизации являются нуклеиновые кислоты или нуклеиновые кислоты, связанные с внутренним вирусным белком. Затем имеет место вторая фаза вирусной репродукции, ведущая к синтезу вирусных компонентов.
Различают три типа взаимодействия вируса с клеткой:
Продуктивный тип — завершается образованием нового поколения вирионов и гибелью (лизисом) зараженных клеток (цитолитическая форма). Некоторые вирусы выходят из клеток, не разрушая их (нецитолитическая форма).
Абортивный тип — не завершается образованием новых вирионов, поскольку инфекционный процесс в клетке прерывается на одном из этапов.
Интегративный тип, или вирогения — характеризуется встраиванием (интеграцией) вирусной ДНК в виде провируса в хромосому клетки и их совместным сосуществованием (совместная репликация).
Вирусные болезни, так же как и другие инфекционные заболевания, возникают при проникновении возбудителя в макроорганизм через определенные входные ворота инфекции. Например, вирусы гриппа, кори и многие другие проникают в организм человека только аэрозольным путем, а вирусы герпеса - множественными путями. Следующий этап вирусной инфекции состоит в адсорбции вируса на рецепторах чувствительных клеток организма хозяина. Однако в отличие от бактериальных, грибковых и протозойных инфекций все последующие события определяются вирусной нуклеиновой кислотой. Это выражается в характерных особенностях, присущих только вирусным инфекциям.
Первая особенность состоит в способности многих РНК- и ДНК-содержащих вирусов вызывать интегративную инфекцию (вирогению), которая происходит при встраивании вирусной нуклеиновой кислоты в хромосому клетки хозяина. Это имеет место при гепатите В, аденовирусной и герпетической инфекции, СПИДе и др. При вирогении отсутствуют стадии репродукции, сборки и выхода вируса из клетки. Клетки с интегрированным вирусным геномом (провирусом) могут сохранить свои функции. Однако в определенных условиях интеграция вирусного генома может привести к мутациям и неконтролируемому делению клетки. Встроенная вирусная ДНК синхронно реплицируется с клеточной ДНК и при делении материнской клетки передается дочерним. При интегративной инфекции вирусный геном может не транскрибироваться или транскрибироваться частично. В случае выщепления из клеточной ДНК происходит его транскрипция и автономная репликация, которая заканчивается выходом вирусного потомства так же, как при продуктивной инфекции.
Вторая особенность обусловлена наличием стадии вирусемии, во время которой вирус циркулирует в крови. В кровь вирус может поступать из лимфатической системы, переноситься лейкоцитами, проникать в кровеносные капилляры из первично инфицированных клеток. Исключение составляют вирусы, распространяющиеся нейрогенным путем (вирусы бешенства, простого герпеса и др.).
Третья особенность заключается в поражении вирусами лимфоцитов - клеток иммунной системы организма человека. Вирусы гриппа, кори, герпеса, полиомиелита, ротавирусы и др. угнетают иммунные реакции Т-лимфоцитов. Вирусы герпеса, вызывающие ветряную оспу и опоясывающий лишай, вирус цитомегалии индуцируют увеличение абсолютного количества Т-лимфоцитов, а вирус клещевого энцефалита вызывает их активацию.
Лимфотропность подавляющего большинства вирусов человека и вотных существенно отражается на патогенезе и исходе вирусных заболеваний, что проявляется в возникновении иммунодефицитных и других иммунопатологических состояний. Наиболее специализированными облигатно-лимфотропными вирусами являются три вируса, поражающие Т-лимфоциты человека, и один - В-лимфоциты. Два первых вируса (HTLV-I и HTLV-II от англ. human T-cell limphotropic virus) вызывают лейкоз вследствие пролиферации Т-лимфоцитов. Третий вирус HTLV-III, или ВИЧ, является возбудителем СПИДа, который в отличие от первых двух вызывает деструкцию Т-лимфоцитов. Вирус герпеса Эпштейна-Баррвоз будитель инфекционного мононуклеоза - вызывает пролиферацию 3-лимфоцитов.
Четвертая особенность, характерная для ряда вирусных инфекций (оспа, бешенство, герпес, корь и др.), состоит в образовании внутриядерных или внутрицитоплазма-тических включений. Они имеют разную форму и величину. Одни из них (базофильные включейия), такие, как тельца Гварниери при оспе и тельца Бабеша-Негри при бешенстве, окрашиваются основными красителями и представляют собой внутриклеточные скопления вируса. Они имеют диагностическое значение. Последствия инфекционного процесса, вызванного вирусами, разнообразны - от сохранения жизнеспособности клетки до широкого спектра поражения. При этом вирусы либо исчезают из организма после выздоровления, либо сохраняются в нем в течение разных сроков, измеряемых в некоторых случаях многими годами. Наличие вируса в организме не всегда сопровождается его выделением. Вирусные болезни протекают в виде продуктивной и персистирующей инфекций. Продуктивная, или острая, инфекция сопровождается репродукцией вирусов в клетках хозяина и сравнительно быстрым выделением их из организма. Острые инфекции можно подразделить на очаговые и генерализованные. Первые проявляются в месте локальной репродукции вируса. Вторые - при генерализации, когда из первичного очага вирус распространяется по организму, главным образом гематогенным путем, формируя вторичные очаги инфекции.
Многие вирусы персистируют в различных клетках организма человека. Например, аденовирусы могут длительно персистировать в миндалинах, вирусы герпеса - в ганглиях тройничного нерва. Пер-систирующая инфекция проявляется в разных формах - латентной, хронической и медленной.
Латентная бессимптомная инфекция характеризуется длительным, а в некоторых случаях пожизненным носительством вируса, который не покидает организм и не выделяется в окружающую среду. В одних случаях это связано с дефектностью вируса, точнее, его генома, в результате чего он утрачивает способность к репродукции и образованию потомства. В других - с интеграцией вирусной ДНК или РНК в клеточный геном и возникновением интегративной инфекции. При этом в клеточную хромосому может встроиться вирусная ДНК полностью, как это имеет место в случае вируса герпеса, либо происходит ее частичное встраивание (вирус гриппа). Последствия такого встраивания трудно предсказать. При герпетической инфекции это часто приводит к возникновению хронической инфекции, при гриппе - не сопровождается видимыми изменениями в течение инфекционного процесса, что может быть связано с неполноценностью вирусной РНК или другими причинами. Они, по-видимому, могут зависеть и от того локуса хромосомы, в который происходит интеграция вирусной нуклеиновой кислоты. В том случае, если она встраивается вблизи промотора, может произойти нарушение регуляции синтеза белка, что приводит к нерегулируемому размножению клеток и возникновению опухоли. Так, например, первичный рак печени у людей, перенесших гепатит, связывают с встраиванием ДНК вируса гепатита В в геном гепатоцитов.
Сайт СТУДОПЕДИЯ проводит ОПРОС! Прими участие :) - нам важно ваше мнение.
Первая особенность состоит в способности некоторых вирусов вызывать интегративную вирусную инфекцию (вирогению), которая происходит при встраивании вирусной нуклеиновой кислоты в хромосому клетки хозяина. Это имеет место при гепатитах В и С, аденовирусах, при герпесе, СПИДе и др. При вирогении отсутствуют такие стадии, как репродукция вируса, сборка и выход из клетки. Клетки с интегрированным вирусным геномом может сохранять свои функции. Однако в определенных условиях интеграция вирусного генома может привести к мутациям и неконтролируемому делению клеток. Встроенный геном вируса синхронно реплицируется с клеточной ДНК и при делении материнской клетки передается дочерним. При интегративной инфекции вирусный геном может не транскибироваться или транскибироваться частично. В случае его выщепления из клеточной ДНК происходит его транскрипция и автономная репликация, которая заканчивается выходом вирусного потомства так же, как при продуктивной инфекции.
Вторая особенностьобусловлена наличием стадии вирусемии, во время которой вирус циркулирует в крови. В кровь вирус может поступать из лимфатической системы, переноситься лейкоцитами, проникать в кровеносные капилляры из первично инфицированных клеток. Исключение составляют вирусы, распространяющиеся нейрогенным путем (вирус бешенства, простого герпеса и др.)
Третья особенностьзаключается в поражении вирусами лимфоцитов – клеток иммунной системы организма человека. Вирусы гриппа, кори, герпеса, полиомиелита, ротавирусы и др. угнетают иммунные реакции Т-лимфоцитов. Вирусы, вызывающие ветряную оспу и опоясывающий лишай, вирус цитомегалии индуцируют увеличение абсолютного количества Т-супрессоров, а вирус клещевого энцефалита вызывает их активацию.
Лимфотропность подавляющего большинства вирусов человека и животных существенно отражается на патогенезе и исходе вирусных заболеваний, что проявляется в возникновении иммунодефицитных и других иммунопатологических состояний. Наиболее специализированными облигатно-лимфотропными вирусами являются три вируса, поражающие Т-лимфоциты человека, и один – В-лимфоциты. Два первых вируса –HTLV-1 и HTLV-II вызывают лейкоз у человека за счет пролиферации Т-лимфоцитов. Третий вирус HTLV- III или ВИЧ, является возбудителем СПИДа, который в отличие от первых двух вызывает декструкцию Т-хелперов. Вирус герпеса Эпстайна-Барра - возбудитель инфекционного мононуклеоза - вызывает пролиферацию В-лимфоцитов.
Четвертая особенностьхарактерна для ряда вирусных инфекций, таких как оспа, бешенство, герпес, корь, состоит в образовании внутриядерных или внутрицитоплазматических включений. Они имеют разную форму и величину. Одни из них являются базофильными включениями, такие как тельца Гварниери при оспе и тельца Бабеша-Негри при бешенстве и представляют собой внутриклеточные скопления вируса. Окрашиваются основными красителями и имеют диагностическое значение.
Последствия инфекционного процесса, вызванного вирусами, разнообразны – от сохранения жизнеспособности клетки до широкого спектра поражения.
Все вирусы обладают цитоцидным действием, а по результатам этих взаимодействий делятся на:
§ Манифестные вирусные инфекции
§ Персистирующие вирусные инфекции
§ Не вызывающие никаких изменений вирусы организма.
Манифестные вирусные инфекции включают все вирусные инфекции, которые протекают с выраженными клиническими проявлениями и свидетельствуют о поражении вирусом тех или иных клеток органов или систем человека и животных. Они протекают в виде продуктивной и персистирующей инфекции.
Продуктивная, или острая инфекция, сопровождается репродукцией вирусов в клетках хозяина и сравнительно быстрым выделением их из организма. Примером данных заболеваний могут служить эпидемии гриппа, полиомиелит, болезнь Боткина, эпидемический паротит, бешенство, клещевой энцефалит, корь и др. Острые вирусные инфекции можно подразделить на очаговые и генерализованные. Очаговые проявляются в месте локальной репродукции вируса. Вторые – при генерализации, когда из первичного очага вирус распространяется по организму, формируя вторичные очаги инфекции.
У иммунокомпетентного хозяина большинство вирусных инфекций, возбудители которых используют первую стратегию паразитизма, самоограничиваются. Смертельные исходы в эпидемических очагах могут достигать очень высоких значений, однако в крупных популяциях людей они никогда не достигают 100%. При недостаточности иммунной системы хозяина, эти же вирусы могут вызывать персистентные или латентные инфекции.
Персистирующая инфекция. Это бессимптомная персистенция вируса, сопровождающаяся его выделением в окружающую среду. Персистирующие вирусные инфекции характеризуются поражением клеток интеграционным провирусом, геном которого редублицируется с клетками хозяина и в комплексе нового качества клетки ведет к ее изменению (механизм близок к взаимодействию с бактериальной клеткой профага). Модус взаимодействия – основа персистирующей инфекции.
В механизме развития вирусной персистирующей инфекции значительная роль отводится особому состоянию возбудителя — дефектным интерферирующим частицам вируса или его мутантам, а также блокированию апоптоза клеток хозяина. Персистирующий инфекционный процесс лежит в основе и проявляется в разных формах – латентной, хронической и медленной инфекции [Гавришева Н.А., Антонова Т.В., 1999].
Латентная инфекция. Это бессимптомная персистенция вируса, не сопровождающаяся его выделением в окружающую среду. При латентной инфекции вирус не удается обнаружить с помощью диагностических приемов в связи с тем, что он находится в дефектной форме или интегрирован в геном клетки. Этот процесс плохо изучен. Значительно большая ясность имеется в вопросе реактивации вируса. Считается, что для этого необходимо действие одного или нескольких активирующих факторов (травма, стресс, иммуносупрессия, суперинфицирование и др.). В результате латентная инфекция переходит манифестную форму — развиваются симптомы, свойственные острой инфекции.
Примерами латентных инфекций служат герпесвирусные (рис.), цитомегаловирусные инфекции, сывороточные гепатиты и ряд других [Гавришева Н.А., Антонова Т.В., 1999].
Схема патогенеза простого герпеса. У большинства лиц с момента первичного инфицирования происходит пожизненная персистенция, находящегося в организме в латентном состоянии вируса. Под воздействием различных активирующих факторов, к которым относятся дефекты иммунной системы, вирус по аксонам выходит из нервных ганглиев, поражая участки кожи и слизистых оболочек, иннервируемые соответствующим нервом. По мере прогрессирования иммуносупрессии активация вируса становится все более частой, в процесс могут вовлекаться все новые ганглии, что приводит к изменению локализации и увеличению распространенности очагов поражения кожи и слизистых оболочек [Гавришева Н.А., Антонова Т.В., 1999]
Однако нельзя полностью согласиться с вышеуказанными авторами, утверждающими, что латентную инфекцию можно рассматривать как фазу инфекционного процесса. Способность вызывать персистирующую или латентную инфекцию не является общим свойством для всех вирусов.
Например, ВНО и отдельные возбудители геморрагических лихорадок либо вызывают смерть инфицированного, либо разрешаются с развитием стерильного иммунитета. Сколь бы многочисленной не осталась пережившая эпидемию популяция, они в ней не сохраняются, а для их поддержания в природе используются другие механизмы.
Поэтому, можно считать, что способность вызывать латентные инфекции является проявлением более глубокой специализации к хозяину вируса, утратившего связь с первичным резервуаром. Латентность этих вирусов необходимо рассматривать только как их свойство, проявляющееся в иммунокомпетентном хозяине, т.е. вирусы такого типа способны использовать обе стратегии паразитизма, но в разной степени. Основную для них вторую стратегию они используют в иммунокомпетентных хозяевах, а в иммунодефицитных вынужденно проявляют первую.
При интегративной инфекции последствия инфекции трудно предсказать. Многое зависит от того локуса хромосомы, в который происходит интеграция вирусной нуклеиновой кислоты. В том случае, если она встраивается вблизи промотора, может произойти нарушение регуляции синтеза белка, что приводит к нерегулируемому размножению клеток и возникновению опухоли. Так, например, первичный рак вируса гепатита В происходит в результате встраивания ДНК вируса гепатита в геном гепатоцитов рядом с промотором.
Хронические вирусные инфекции также рассматривают как одну из форм персистенции вируса, которая продолжается в течение нескольких месяцев и лет. Данную форму инфекции вызывают аденовирусы, вирусы гепатита, герпеса и др., которые периодически выделяются из организма больного во внешнюю среду.
Интеграция РНК вирусов в клеточный геном происходит путем обратной транскрипции. В 80- годах в клетках людей были обнаружены тысячи копий ретровирусных генов неизвестного происхождения, напоминающие транспозоны бактерий. Их называют ретротранспозонами вследствие их образования путем обратной транскрипции. Встраивание этих ретротранспозонов в хромосомы клеток людей и животных, так же как и в клетки бактерий, дрожжей и насекомых, происходит беспорядочно. Это может привести к мутациям, нарушающим работу мутировавшего гена, или изменению уровня его экспрессии (активации). Таким образом, данный ген становится онкогеном, индуцирующим образование опухоли. У человека продукты обратной транскрипции (ретротранспозоны, эндогенные провирусы) составляют 10% клеточного генома.
Для поддержания в природе возбудителей инфекций, использующих стратегию паразитирования II типа, размер инфицированной популяции имеет значительно меньшее значение, чем для возбудителей, использующих стратегию I типа.
Медленные инфекции. Их вызывают паразитические микроорганизмы, использующие только вторую стратегию паразитизма, т.е. уклонение от иммунной системы хозяина. Медленные инфекции характеризуются длительным инкубационным периодом, прогрессирующим течением и неизбежной смертью. Кроме вирусов, медленные инфекции вызывают прионы — инфекционные безнуклеиновые структуры, состоящие из низкомолекулярного белка, молекулярная масса которого составляет 27. 30 кд.
Прионы имеют свойства, типичные для обычных вирусов, однако отличаются от них выраженной устойчивостью к действию многих физических факторов. Они не имеют сердцевины из нуклеиновых кислот, окруженных протеинами, а также липидной оболочки, и в связи с этим при электронной микроскопии не распознаются как вирусы. Медленные инфекции могут вызывать некоторые вирусы, использующие первую стратегию паразитизма (например, вирусы кори — подострый склерозирующий панэнцефалит, клещевого энцефалита — прогредиентная форма клещевого энцефалита). Прогредиентная форма клещевого энцефалита сопровождается спонгиозной дегенерацией астроцитарной глии и глиосклерозом. К медленным инфекциям относятся: ВИЧ- инфекции, Т-клеточная лимфома, подострый герпетический энцефалит и др. Но это не является всеобщей закономерностью и происходит только при определенном состоянии иммунной системы, когда эффективные иммунные реакции макроорганизма на вирус заблокированы.
Ничего не вызывающие вирусы организма выделяются в достаточном количестве с поверхности слущивающегося эпидермиса, из испражнений, отделяемого слизистых, а также из окружающей среды. Определение содержания энтеровирусов в питьевой воде имеет на сегодня большое эпидемическое значение в санитарной вирусологии.
Резервуаром вирусных инфекций энцефалита, геморрагических и других видов лихорадок являются лани, олени, мыши, а переносчиками этих вирусных инфекций чаще всего являются клещи, получившие вирус от носителя, причем некоторые виды вирусных инфекций передаются трансовариально потомству клещей. Человек, попавший в это звено и укушенный клещом, заболевает одним из видов уже манифестной вирусной инфекции (клещевой энцефалит, гемморрагическая лихорадка и др.).
| | | следующая лекция ==> | |
| Особенности вирусных инфекций. Вирусы являются облигатными паразитами, использующими генетические ресурсы хозяина в значительно большей степени | | | Понятие об эпидемическом процессе |
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Взаимодействие вируса с клеткой хозяина - это сложный «многоступенчатый процесс, который начинается с адсорбции вирусных частиц на рецепторах клетки хозяина и продолжается после их проникновения внутрь клетки. В результате такого взаимодействия мовивается либо продуктивная, либо абортивная, либо интегративная форма клеточной инфекции. При продуктивной форме происходит размножение, точнее, репродукция (лат. reproduce - воспроизводить) вируса, при абортивной - ее нарушение на одном из этапов, при интегративной - интеграция вирусной нуклеиновой кислоты в клеточный геном.
Продуктивная инфекция. Репродукция вирусов.
В 50-х годах было установлено, что размножение, или репродукция, вирусов происходит путем репликации их нуклеиновой кислоты и биосинтеза белков с последующей самосборкой вириона. Этот процесс происходит в разных частях клетки - ядре или цитоплазме, вследствие чего получил название дизъюнктивного, т.е. разобщенного размножения.
Вирусная репродукция представляет собой уникальную форму выражения чужеродной (вирусной) информации в клетках человека и животных, насекомых, растений и бактерий, которая состоит в подчинении клеточных матрично-генетических механизмов вирусной информации.
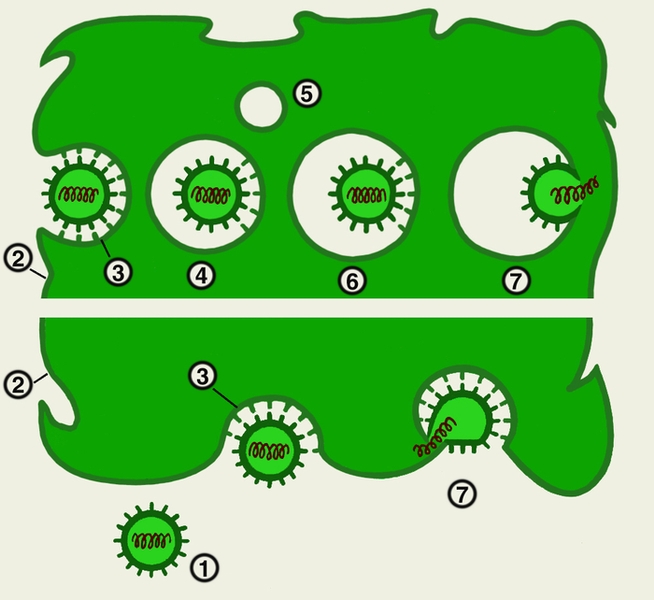
Схема проникновения вирусов в клетку путем рецепторного эндоцитоза (вверху) и через плазматическую мембрану клетки (внизу):
1 — вирусная частица;
2 — плазматическая мембрана клетки;
3 — ямка на поверхности клетки, содержащая рецепторы;
4 — образующаяся клеточная вакуоль;
5 — сливающаяся клеточная вакуоль;
6 — клеточная вакуоль, образующаяся после слияния (рецептосома);
7 — выход генетического материала вируса в цитоплазму.
1-я стадия. Взаимодействие вируса с клеткой начинается с процесса адсорбции, т. е. прикрепления вирусов к поверхности клетки. Это высокоспецифический процесс. Вирус адсорбируется на определенных участках клеточной мембраны — так называемых рецепторах. Клеточные рецепторы могут иметь разную химическую природу, представляя собой белки, углеводные компоненты белков и липидов, липиды. Число специфических рецепторов на поверхности одной клетки колеблется от 10 4 до 10 5 . Следоват ельно, видоспецифичны — вирусы архибактерий не могут инфицировать клетку E. coli, а многие колифаги не проникают в клетку шигеллы. Наиболее выражена тканевая специфичность вирусов животных и человека. Так, вирусы гепатитов поражают гепатоциты, вирус Эпштейна-Барр (вызывает инфекционный мононуклеоз) обладает тропностью к B-лимфоцитам, ВИЧ — к T-лимфоцитам, кишечные вирусы — к энтероцитам, кардиотропностью обладают вирусы Коксаки B. Целый ряд вирусов обладает тропностью не к одному, а к нескольким типам клеток. Так полиовирусы тропны к клеткам респираторного тракта, желудочно-кишечного тракта (ЖКТ), центральной нервной системы (ЦНС). Вирус гепатита C (ВГC) лимфотропен и гепатотропен.
Специфическое сродство вирусов к клеткам и тканям определяется двумя механизмами:
— присутствием на клеточной поверхности специфических для вируса рецепторов.
— содержанием в системе активирующих ферментов, необходимых для протеолитического расщепления вирусных поверхностных белков и проявления инфекционной активности вируса.
2-я стадия - проникновение вируса в клетку хозяина - происходит несколькими путями.
Рецепторно-опосредованный эндоцитоз характеризуется образованием в месте взаимодействия вириона с клеточным рецептором окаймленных пузырьков, в формировании которых принимают участие белки-клатрины.
Виропексис. Этим путем в клетку проникают сложноустроенные вирусы. Он заключается в слиянии мембран - вирусного суперкапсида с клеточной или ядерной мембраной. Данный процесс происходит при помощи специального белка слияния - F-белка, который находится в суперкапсиде. В результате виропексиса капсид оказывается в клетке хозяина, а суперкапсид вместе с белком встраивается в ее плазматическую мембрану (вследствие чего клетка приобретает способность сливаться с другими клетками, что приводит к передаче вируса этим клеткам).
Фагоцитоз. Данным путем вирус проникает в фагоцитирующие клетки, что приводит к незавершенному фагоцитозу.
Проникновение вируса в организм хозяина у разных биологических видов решается по-разному.
1. Вирусы растений проникают в организм хозяина по типу раневых инфекций, где распространяются по плазмодесмам, ксилеме и флоэме.
2. Вирусы бактерий — путем введения нуклеиновой кислоты в тело клетки или путем проникновения вириона.
3. Вирусы насекомых попадают в организм хозяина в процессе питания или размножения.
4. Вирусы животных и человека при инфицировании организма хозяина проходят более сложный путь. Одни вирусы (вирус гриппа, ротавирусы) реплицируются и вызывают заболевание в месте проникновения в организм (входные ворота инфекции). Другие вирусы, попав в организм хозяина с использованием того или иного механизма, проходят стадию распространения.
3-я стадия - транспорт вируса внутри клетки. Он происходит с помощью внутриклеточных мембранных пузырьков, в которых вирус переносится на рибосомы, эндоплазматическую сеть или в ядро.
Распространение вируса в организме сопровождается виремией (вирусемией) — циркуляцией вируса в крови, что свидетельствует о генерализации инфекции.
Различают несколько путей распространения вирусов в организме:
1. Нейронный путь (вирусы бешенства, герпеса).
2. Лимфатический путь (реовирусы, полиомавирусы).
3. Гематогенный путь, ассоциированный с клеточными компонентами и плазмой крови (вирус краснухи, вирусы гепатита B и C, цитомегаловирус, энтеровирусы).
Сохранение вируса как биологического вида обеспечивает его восприимчивый хозяин, который является основным элементом экологической ниши вируса. Способность клеток или организма хозяина заражаться называется восприимчивостью.
5-я стадия называется эклипс-фазой, которая характеризуется исчезновением вириона, поскольку он перестает обнаруживаться при электронной микроскопии. В эту стадию начинается синтез компонентов вириона, т.е. его репродукция. Она носит раздельный характер, поскольку компоненты вириона синтезируются в разных частях клетки: белки на рибосомах, нуклеиновые кислоты в ядре или цитоплазме. Вирус использует для этого генетический аппарат клетки, подавляя необходимые ей самой синтетические реакции.
Эта стадия начинается с транскрипции и репликации вирусного генома. Транскрипция вирусного генома двунитевых ДНК-содержащих вирусов происходит, так же как и клеточного генома, по триаде ДНК- иРНК- белок. Различия касаются только происхождения фермента ДНК-зависимой РНК-полимеразы, необходимой для данного процесса. У вирусов, геном которых транскрибируется в цитоплазме клетки хозяина (например, вирус оспы), имеется собственная вирусспецифическая РНК-полимераза. Вирусы, геномы которых транскрибируются в ядре (папова- и аденовирусы, вирусы герпеса), используют содержащуюся там клеточную РНК-полимеразу II или III.
У РНК-содержащих вирусов транскрипция их генома осуществляется несколькими путями:
1. Вирусы с негативным геномом (минус-нитевые), к которым относятся орто-, парамиксо- и рабдовирусы, имеют в своем составе вирусспецифическую РНК-полимеразу или транскриптазу. Они синтезируют иРНК на матрице геномной РНК. Подобный фермент отсутствует в нормальных клетках, но синтезируется клетками, зараженными вирусами. Он находится в составе как однонитевых, так и двунитевых РНК-содержащих вирусов.
2. У вирусов с положительным геномом (плюс-нитевые), к которым относятся пикорна-, тогавирусы и др., функцию иРНК выполняет сам геном, который транслирует содержащуюся в нем информацию на рибосомы клетки хозяина.
3. Особняком стоит группа РНК-содержащих ретровирусов, в составе которых имеется обратная транскриптаза, или ревертаза. Уникальность этого фермента состоит в его способности переписывать информацию с РНК на ДНК. Этот процесс называется обратной транскрипции.
Как отмечалось выше, количество генов в вирусном геноме весьма ограничено. Поэтому для увеличения количества вирусной информации существует своеобразный трансляционный механизм, функционирующий через иРНК, который передает значительно больше информации, чем записано в вирусной нуклеиновой кислоте. Это достигается разными путями, например при транскрипции информации с переписывающихся участков ДНК на иРНК путем сплайсинга (вырезание бессмысленных кодонов и сшивание концов), а также при считывании антикодонами тРНК одной и той же молекулы иРНК с разных нуклеотидов. При этом образуются новые триплеты, увеличивающие количество транслируемой информации.
Вирус-специфическая информация транслируется на рибосомы клетки хозяина, которые предварительно освобождаются от клеточных белков и собираются в вирус-специфические полисомы.
6-я стадия - сборка вириона - состоит прежде всего в образовании нуклеокапсидов. Поскольку синтез вирусных нуклеиновых кислот и белков в клетке происходит в разных ее структурах, необходима транспортировка составных частей вириона в одно место сборки. При этом вирусные белки и нуклеиновые кислоты обладают способностью узнавать и самопроизвольно соединяться друг с другом. В основе самосборки простых вирионов лежит способность вирусных полипептидов соединяться в капсомеры, которые, располагаясь вокруг осей симметрии, образуют многогранник. В других случаях полипептиды в виде спирали окружают вирусную нуклеиновую кислоту.
Многие простые вирионы собираются на репликативных комплексах мембранах эндоплазматического ретикулума. У сложных вирионов сборка нуклеокапсида начинается на репликативных комплексах, а затем продолжается на плазматической мембране, с наружной стороны которой располагаются суперкапсидные гликопротеиды. Затем гликопротеидные и примыкающие к ним с другой стороны нуклеокапсидные участки выпячиваются через клеточную мембрану, образуя почку, как это имеет место у орто- и парамиксовирусов, рабдовирусов. После отделения почки, содержащей нуклеокапсид и суперкапсидные белки, образуются свободные вирионы. Они либо через клеточную плазматическую мембрану проходят во внеклеточное пространство, либо через мембрану эндоплазматического ретикулума проникают в вакуоль эндоплазматической сети. При этом мембранные липиды обволакивают почку, вытесняя из нее белки. Многие ДНК-содержащие вирусы, например вирус герпеса, собираются в ядре клетки на ее мембране, где образуются нуклеокапсиды. Затем они отпочковываются в перинуклеарное пространство, приобретая внешнюю оболочку. Дальнейшее формирование вириона происходит в мембранах цитоплазматического ретикулума и в аппарате Гольджи, откуда вирус транспортируется на поверхность клетки.
Существуют следующие общие принципы сборки вирусов, имеющих разную структуру:
1. формирование вирусов является многоступенчатым процессом с образованием промежуточных форм;
2. сборка просто устроенных вирусов заключается во взаимодействии молекул вирусных нуклеиновых кислот с капсидными белками и образовании нуклеокапсидов (например, вирусы полиомиелита). У сложно устроенных вирусов сначала формируются нуклеокапсиды, с которыми взаимодействуют белки суперкапсидных оболочек (например, вирусы гриппа);
3. формирование вирусов происходит не во внутриклеточной жидкости, а на ядерных или цитоплазматических мембранах клетки;
4. сложно организованные вирусы в процессе формирования включают в свой состав компоненты клетки-хозяина (липиды, углеводы).
7-я стадия - выход вирусных частиц из клетки - происходит двумя путями.
Первый тип — взрывной — характеризуется одновременным выходом большого количества вирусов. При этом клетка быстро погибает. Такой способ выхода характерен для вирусов, не имеющих суперкапсидной оболочки.
Время, необходимое для осуществления полного цикла репродукции вирусов, варьирует от 5—6 часов (вирусы гриппа, натуральной оспы и др.) до нескольких суток (вирусы кори, аденовирусы и др.). Образовавшиеся вирусы способны инфицировать новые клетки и проходить в них указанный выше цикл репродукции.
Простые вирусы, лишенные суперкапсида, например пикорнавирусы, аденовирусы и др., вызывают деструкцию клетки и попадают во внеклеточное пространство. Другие вирусы, имеющие липопротеидную внешнюю оболочку, выходят из клетки путем почкования, в результате чего в течение длительного времени она сохраняет свою жизнеспособность. Такой путь характерен для вируса гриппа и др.
Интегративная инфекция. Интеграция (встраивание) вирусной нуклеиновой кислоты в клеточный геном.
Данный путь взаимодействия между вирусом и клеткой хороша не одинаков для ДНК- и РНК-содержащих вирусов. В первом случае вирусная ДНК в кольцевой форме интегрирует в клеточный геном. При этом место интеграции определяется гомологичными нуклеотидными последовательностями, имеющимися в определенных участках - ДНК сайтах при участии ряда ферментов: рестриктаз, эндонуклеаз, лигаз. Вирус, интегрированный в клеточный геном, называют провирусом.
Провирус может реплицироваться в составе клеточного генома пропорционально делению клетки. При этом каждая дочерняя клетка получает копию провирусного генома. В другом случае амплификация провирусной ДНК с увеличением числа копий провируса без его выщепления из клеточного генома может привести к встраиванию провируса в другую хромосому. Выщепление провируса из клеточного генома и его проникновение в новую клетку может вызвать продуктивную инфекцию.
В случае РНК-содержащих вирусов включение РНК в клеточный геном происходит путем обратной транскрипции. Механизм обратной транскрипции состоит в первоначальном образовании ДНК-транскрипта на матрице РНК при обязательном участии обратной транскриптазы. Этот транскрипт представляет собой одну нить ДНК, являющуюся матрицей для образования второй нити. Затем образовавшийся двунитевои ДНК-транскрипт замыкается в кольцо и встраивается в клеточный геном. Данный процесс объединения вирусной нуклеиновой кислоты с хромосомой клетки хозяина называется вирогениеи. В интегрированном состоянии вирусная ДНК может транскрибироваться в составе клеточного генома при участии клеточных РНК-полимераз.
Биологический смысл интегративного типа взаимодействия между вирусом и клеткой хозяина можно видеть прежде всего в сохранении вирусной информации в составе клеточного генома и ее передаче потомству. Вместе с тем это в определенной степени отражается и на эволюции некоторых вирусов (например, бактериофагов), которые при выщеплении из состава клеточной хромосомы могут захватывать отдельные ее гены.
С другой стороны, подобный тип взаимодействия может отразиться на судьбе клеток хозяина в зависимости от расположения локуса, в котором происходит интеграция вирусного генома, вплоть до расстройства регуляции синтеза белка и неконтролируемого деления клетки. Это может привести к онкогенной трансформации клеток хозяина и развитию разнообразных опухолей.
Дефектные вирусы.
Дефектные вирусы - вирусы, утратившие в процессе репродукции часть своего генома:
- дефектные интерферирующие частицы представляют собой вирионы, содержащие только часть генетической информации исходного вируса; репродуцируются только при участии родственного им вируса-помощника;
- вирусы-спутники отличаются для своей репродукции требуют участия любого вируса-помощника, не обязательно родственного исходному вирусу. например, вирус гепатита D (дельта) репродуцируется только в присутствии вируса гепатита В;
- интегрированные геномы представляют собой провирусы, т. е. вирусные геномы, встроенные (интегрированные) в хромосому клетки хозяина при интегрированной инфекции, которые потеряли способность превращаться в полноценный вирус;
- псевдовирионы - вирионы, имеющие нормальный капсид, содержащий часть собственной нуклеиновой кислоты и фрагменты нуклеиновой кислоты своего хозяина, либо часть хромосомы клетки хозяина и часть нуклеиновой кислоты другого вируса.
Значение дефектных вирусов состоит в их способности переносить генетический материал из клетки донора в клетку реципиента.
Читайте также:
