Все известные на данный момент ви́русы архе́й имеют геномы, представленные ДНК: одноцепочечной или двухцепочечной, кольцевой [en] или линейной. Недавно, однако, в горячих источниках Йеллоустонского национального парка, которые населены почти исключительно видом архей Sulfolobus solfataricus [en] , с помощью метагеномики обнаружили вирусный РНК-геном, отдалённо напоминающий эукариотические РНК-вирусы [en] * , поэтому, возможно, существуют и РНК-содержащие вирусы архей [1] .
По состоянию на 2019 год вирусы архей представлены 17 семействами. Стоит отметить, что вирусы архей составляют две сильно различающиеся группы. К первой относят вирусы, которые структурно и генетически близки к вирусам бактерий и эукариот, а ко второй — вирусы, уникальные для архей и мало похожие на вирусы других клеточных форм. Практически все специфические для архей вирусы поражают представителей типа Crenarchaeota, а вирусы, близкие к бактериофагам и вирусам эукариот, чаще всего паразитируют на археях типа Euryarchaeota [1] .
Содержание
Строение вирионов [ | код ]
Вирусы, специфичные для архей, нередко имеют вирионы необычной формы. Так, представители семейства Ampullaviridae [en] имеют вирионы в виде бутылок из-под шампанского, причем поверх белков капсида они покрыты липидной оболочкой. У членов семейства Spiraviridae [en] вирионы имеют вид спиралей. Столь необычная форма вирионов у вирусов этих двух семейств связана с особым способом упаковки генома при помощи белков капсида [1] .
Некоторые специфичные для архей вирусы имеют веретеновидные капсиды. Среди них у вирусов семейства Fuselloviridae [en] на одном из заострённых концов капсида находится пучок белковых филаментов, а у членов семейства Bicaudaviridae [en] на одном или двух концах капсида находятся одиночные придатки в виде хвостов. Не менее необычна морфология вирионов Guttaviridae [en] : у этих вирусов вирионы тоже похожи на веретено, однако один их конец закруглён и имеет каплевидную форму [1] .
У многих специфичных для архей вирусов имеются нитчатые вирионы, которые могут нести специальные придатки, предназначенные для распознавания клеток архей. Иногда, как у вирусов семейства Tristromaviridae, капсид сформирован не одним, а тремя типами белков [1] .
Некоторые вирусы, специфичные для архей, имеют сферические вирионы, причём иногда поверх капсида находятся липидная мембрана и ещё один слой белков, из-за чего частицы принимают икосаэдрическую форму [1] .
Вирионы Pleolipoviridae [de] по строению похожи на везикулы, которые образуют многие археи: голая геномная ДНК находится внутри мембранного пузырька, который пронизан белками двух типов. Такие пузырьки могут содержать как одноцепочечную, так и двухцепочечную ДНК линейной или кольцевой формы [1] .
Вирионы некоторых вирусов архей удалось не только рассмотреть под электронным микроскопом, но и детально изучить с помощью криоэлектронной микроскопии. Так, было обнаружено, что геномы некоторых вирусов архей в капсидах находятся в А-форме. Это первый известный случай, когда А-форма ДНК присутствует в живых организмах в нормальных условиях. Один из вирусов с геномом в виде А-ДНК, AFV1, имеет очень тонкую липидную оболочку с необычным химическим составом — главным её компонентом является липид глицеролдибифитанилглицеролтетраэфир (GDGT-0), имеющий необычную U-образную конфигурацию. В мембране, состоящей их таких липидов, их гидрофильные головки обращены наружу, а гидрофобные дуги — внутрь. Наряду с фосфолипидным бислоем и монослоем архей такое строение можно считать третьим из известных типов биологических мембран [1] .
Геномы [ | код ]
Все выделенные на данный момент вирусы архей имеют ДНК-геномы (хотя, как отмечалось выше, в горячих источниках Йеллоустона с помощью метагеномики удалось найти РНК-геном возможного вируса архей). В большинстве случаев геном представлен двухцепочечной молекулой ДНК, и лишь у членов семейств Spiraviridae и Pleolipoviridae геномы состоят из одноцепочечной ДНК. Размеры геномов архейных вирусов варьируют от 5300 пар оснований (п. о.) у вируса APBV1 (это один из мельчайших известных геномов ДНК-содержащих вирусов) до 143 800 п. о. у миовируса HGTV-1. Как правило, вирусы, специфичные для архей, имеют меньшие геномы, чем вирусы архей, родственные бактериофагам и вирусам эукариот [2] .
Механизмы репликации геномов вирусов архей экспериментально изучены лишь для небольшого числа вирусов. Известно, что ДНК представителей порядка Caudovirales (которые, кстати, имеют самые большие геномы среди архейных вирусов) кодирует часть или даже все компоненты аппарата репликации ДНК: ДНК-полимеразы, белки скользящего зажима ( PCNA [en] ), праймазы и хеликазы. Имеющие более скромные размеры геномов вирусы архей, как правило, кодируют белки, которые необходимы для привлечения аппарата репликации клетки-хозяина. Стоит, однако, отметить, что в геномах многих вирусов, специфичных для архей, не удалось найти белки, связанные с репликацией ДНК, так что они либо полностью зависимы от аппарата репликации клетки-хозяина, либо используют уникальные, пока ещё не изученные механизмы репликации ДНК. Например, удалось показать, что и инициация, и терминация репликации генома липотриксвируса AFV1 связаны с рекомбинационными процессами [1] .
Механизмы упаковки генома в капсид у вирусов архей детально не изучены. Тем не менее известно, что члены порядка Caudovirales имеют гомологи терминазы, которая упаковывает геномную ДНК в пустой капсид. Механизм формирования вирионов с использованием терминазы используют также вирусы бактерий и эукариот. Можно предположить, что упаковка генома в капсид у вирусов архей протекает так же, как и у бактериофагов и вирусов эукариот, но в плане репликации ДНК вирусы архей или целиком зависят от клетки-хозяина, либо используют уникальные, пока ещё не изученные механизмы [1] .
Взаимодействие с клеткой-хозяином [ | код ]
Выход зрелых вирионов из клетки архей во многих случаях напоминает отпочковывание от эукариотической клетки-хозяина у вируса гриппа, ВИЧ и вируса Эбола. Когда вирион покидает клетку археи, он забирает с собой фрагмент её мембраны, который становится дополнительной оболочкой, лежащей поверх капсида. У некоторых вирусов архей финальные стадии созревания вирионов происходят уже после выхода из клетки, когда капсид претерпевает морфологические перестройки [1] .
У некоторых вирусов архей (представителей семейств Rudiviridae [en] и Turriviridae [en] ) все стадии созревания вириона проходят в цитоплазме клетки. Новые вирусные частицы покидают клетку через специальные структуры с семиосевой симметрией на их поверхности, которые получили название вирусассоциированных пирамид (virus-associated pyramids, VAP). VAP образуются на внутренней поверхности мембраны заражённой клетки, проходят сквозь её поверхностный S-слой и открываются на финальных этапах инфекции, давая возможность вирионам выйти из клетки [1] .
Выход некоторых вирусов архей из клетки сопровождается её лизисом. К их числу относятся вирусы семейства Tristromaviridae, которые, хотя и развиваются целиком в цитоплазме, каким-то образом приобретают липидную оболочку. Примечательно, что вирус ψM2 кодирует фермент псевдомуреинэндоизопептидазу, который разрушает псевдомуреиновую [en] клеточную стенку археи [1] .
Эволюция и родственные связи [ | код ]
Вирусы, специфичные для архей, как правило, заражают только представителей типа Crenarchaeota. Они отличаются от всех других вирусов не только нестандартной морфологией вирионов, но и генетически: около 90 % их генов не имеет гомологов в существующих базах данных. В геномах некоторых вирусов архей не удалось найти ни одного белка, для которого существовал бы функционально охарактеризованный белок-гомолог [1] .
Иногда, если гомологию не удаётся установить по нуклеотидным или аминокислотным последовательностям, на помощь приходят пространственные структуры. Действительно, для различных белков вирусов архей получили пространственные структуры, но ситуация от этого не стала яснее: оказалось, что во многих из них содержатся совершенно уникальные структурные мотивы. Более того, функции многих генов вирусов архей совершенно непонятны: так, оказалось, что вирус SSV1 выживает без половины своих генов. Можно предположить, что такие неохарактеризованные гены-сироты кодируют белки, участвующие во взаимодействии вируса с клеткой архей, например, противодействующие системам CRISPR-Cas [1] .
Впрочем, многие вирусы архей родственны некоторым бактериофагам и вирусам эукариот. Однако вирусы, специфичные для архей, стоят особняком среди всех ДНК-содержащих вирусов. Более того, различные группы вирусов, специфичных для архей, неродственны друг другу и эволюционируют независимо друг от друга. Высказывается предположение, что некоторые группы специфичных для архей вирусов появились на заре эволюции клеточной жизни и были впоследствии утрачены бактериями и эукариотами. Другие группы специфичных архейных вирусов могли появиться в момент обособления домена архей или даже позже, в отдельных группах архей [1] .
Научный интерес представляет родство некоторых вирусов архей — и лишённых капсидов мобильных генетических элементов (например, плазмид). Эти вирусы, как и мобильные генетические элементы, имеют родственные гены основных белков репликативного аппарата [1] [2] .
Когда у нас затягиваются царапина на коже, растут волосы и ногти и происходят всякие другие процессы обновления тканей, клетки нашего тела делятся. Перед началом этого процесса в них происходит копирование хромосом, чтобы при делении каждая из дочерних клеток получила свою порцию генетической информации. Для этого спирали ДНК, из которых состоят хромосомы, расплетаются и к каждой из получившихся нитей достраивается новая. Этот процесс начинается не где попало, а с особой последовательности генов, называемой точкой начала репликации. Такие точки есть у всех живых организмов, они позволяют регулировать клеточный цикл и следить за тем, чтобы копирование хромосом происходило одновременно.
Торстен Аллерс (Thorsten Allers) из университета Ноттингема (Великобритания) и его коллеги поставили опыт: они удалили точки начала репликации на хромосомах у одного из видов архей, галофераксов — Haloferax volcanii — и посмотрели, что из этого выйдет.
![]()
Живые существа делятся на три домена: эукариоты (мы относимся именно к ним), бактерии и археи. Археи знамениты своей способностью выживать в экстремальных условиях, таких как горячие источники или соленые озера. Галофераксы, например, обитают в Мертвом море.
Бардак в геноме
Геном у представителей трех доменов организован по-разному. Наиболее сложным образом — у эукариот. Например, человеческий геном разбит на 23 хромосомы, каждая из которых представлена в клетке в двух вариантах: по одной от папы и от мамы. Чтобы при делении дочерние клетки получили всю необходимую генетическую информацию, копирование всех 23 пар хромосом должно начинаться одновременно. Для этого и нужны точки начала репликации: если копирование ДНК начинается в строго определенных местах, клетке проще контролировать и время старта. На хромосомах человека точек начала репликации тысячи, если одна почему-либо не сработает, процесс может быть начат в другой точке.
У бактерий этот процесс выглядит проще: у них весь геном умещается в одной хромосоме, а ее копий в клетке может быть от одной до четырех. На каждой хромосоме есть одна точка начала репликации, и копирование ДНК всегда идет в одном направлении. Это позволяет бактериям размножаться очень быстро, например, постоянный объект всевозможных биологических экспериментов, кишечная палочка, делится раз в 25 минут.
Смертельный номер
Все мы немножко. вирусы?!
Однако это вызывает следующий вопрос: зачем галофераксу вообще нужны эти точки, если без них ему не только хорошо, но и даже лучше? Аллерс полагает, что самые первые клетки имели множество копий генома, как галоферакс, и при его копировании полагались на механизм рекомбинации.
С точками начала репликации древние клетки стали делиться медленнее, потому что копирование ДНК стало возможно начинать только в одной точке, но зато эту точку оказалось удобно использовать для регуляции клеточного цикла, чтобы контролировать время и место начала репликации. А если репликацию можно начинать в определенное время, то можно попытаться снова ускорить процесс, разбив геном на небольшие хромосомы, копирование которых будет начинаться в одно и то же время.
Возможно, этому поспособствовало то, что вместе с точками начала репликации древние клетки унаследовали от вирусов и другие механизмы, помогающие копировать ДНК и регулировать клеточный цикл. Более того, есть гипотеза, согласно которой самые первые клетки использовали для хранения и передачи наследственной информации молекулы РНК, а не ДНК. ДНК в клетки могли принести вирусы, и в какой-то момент репликационный механизм переключился с клеточной РНК на вирусную ДНК, вместе с которой клетками могли быть унаследованы и точки начала репликации.
Чем галоферакс похож на раковые клетки
Практическое применение открытия Аллерса и его коллег может лежать в области изучения раковых клеток. Галоферакс и раковые клетки — это далеко не одно и то же, однако некоторые сходства у них есть. В частности, для раковых клеток характерно большое количество копий генома и использование механизма рекомбинации, которое делает геном клетки очень нестабильным.
Екатерина Боровикова
Вирусы заражают все клеточные формы жизни, и археи — не исключение. Хотя сейчас известно гораздо меньше вирусов архей, чем вирусов бактерий и эукариот, разнообразие устройства их вирионов и используемых молекулярных механизмов поражает воображение: некоторые из них имеют вирионы в форме бутылки, у других генетический материал в вирионе хранится в форме А-ДНК, третьи обзавелись уникальным типом биологических мембран или используют для выхода из зараженной археи ранее неизвестные клеточные структуры. Наша статья посвящена этим пока еще малоизученным, но удивительно самобытным генетическим элементам.
Давно прошли те времена, когда археи считались обитателями исключительно тех мест, где никакая другая клеточная жизнь существовать не может, например, горячих источников. В действительности, архей можно найти везде, даже в желудочно-кишечном тракте человека, однако наиболее многочисленны они на глубоководьях морей и океанов. Так, показано, что археи, окисляющие ионы аммония до нитрата (они относятся к типу Thaumarchaeota ) являются одной из самых многочисленных клеточных форм жизни в океанах. Неудивительно, что и вирусы архей также очень многочисленны: метагеномный анализ показал, что в некоторых зонах на долю вирусов архей приходится около 10% самых распространенных там вирусов. Объемы клеток архей, которые погибают из-за вирусов, тоже впечатляют: установлено, что в верхних 50 см океанического донного грунта клетки архей, лизированные вирусами, составляют до трети всей ежегодной биомассы погибших микробов, что соответствует 0,3–0,5 гигатоннам углерода ежегодно [1] .
Все известные на данный момент вирусы архей имеют геномы, представленные ДНК: одноцепочечной или двухцепочечной, кольцевой или линейной. Недавно, однако, в горячих источниках Йеллоустонского национального парка, которые населены почти исключительно археей Sulfolobus solfataricus , с помощью метагеномики обнаружили вирусный РНК-геном, отдаленно напоминающий эукариотические РНК-вирусы, поэтому, возможно, существуют и РНК-содержащие вирусы архей.
Пока вирусы архей представлены 17 семействами. Стоит отметить, что вирусы архей, вообще-то, составляют две сильно различающиеся группы. К первой относят вирусы, которые структурно и генетически близки к вирусам бактерий и эукариот, а ко второй — вирусы, уникальные для архей и мало похожие на вирусы других клеточных форм. Практически все специфические для архей вирусы поражают представителей типа Crenarchaeota , а вирусы, близкие к бактериофагам и вирусам эукариот, чаще всего паразитируют на археях типа Euryarchaeota [1] .
Строение вирусных частиц
Вирусы, специфичные для архей, нередко имеют вирионы необычной формы (рис. 1 а ).
Так, представители семейства Ampullaviridae имеют вирионы в виде бутылок из-под шампанского, причем поверх белков капсида они обтянуты липидной оболочкой. А у членов семейства Spiraviridae вирионы в виде спиралей. Столь необычная форма вирионов у вирусов этих двух семейств связана с особым способом упаковки генома при помощи белков капсида [1] .
ся липидная мембрана и еще один слой белков, из-за чего частицы принимают икосаэдрическую форму [1] .
Вирионы Pleolipoviridae по строению похожи на везикулы, которые образуют многие археи: голая геномная ДНК находится внутри мембранного пузырька, который пронизан белками двух типов. Такие пузырьки могут содержать как одноцепочечную, так и двухцепочечную ДНК линейной или кольцевой формы [1] .
Чем можно объяснить столь необычную морфологию вирусов архей? Вирионы некоторых из них удалось не только рассмотреть под электронным микроскопом, но и детально изучить с помощью криоэлектронной микроскопии. Так ученые выяснили, что геномы некоторых вирусов архей в капсидах находятся в А-форме ! Это первый известный случай, когда А-форма ДНК присутствует в живых организмах в нормальных условиях. Один из вирусов с геномом в виде А-ДНК, AFV1, имеет очень тонкую липидную оболочку с необычным химическим составом — главным ее компонентом является липид глицеролдибифитанилглицеролтетраэфир (GDGT-0), имеющий необычную U-образную конфигурацию (рис. 3). В мембране, состоящей их таких липидов, их гидрофильные головки обращены наружу, а гидрофобные дуги — внутрь. Наряду с фосфолипидным бислоем и монослоем архей такое строение можно считать третьим из известных типов биологических мембран [1] .
Геномы
Все выделенные на данный момент вирусы архей имеют ДНК-геномы (хотя, как отмечалось выше, в горячих источниках Йеллоустона с помощью метагеномики удалось найти РНК-геном возможного вируса архей). В большинстве случаев геном представлен двухцепочечной молекулой ДНК, и лишь у членов семейств Spiraviridae и Pleolipoviridae геномы состоят из одноцепочечной ДНК. Размеры геномов архейных вирусов варьируют от 5300 пар нуклеотидов (п.н.) у клававируса APBV1 (это один из мельчайших известных геномов ДНК-содержащих вирусов) до 143 800 п.н. у миовируса HGTV-1. Как правило, вирусы, специфичные для архей, имеют меньшие геномы, чем вирусы архей, родственные бактериофагам и вирусам эукариот [2] .
Механизмы репликации геномов вирусов архей экспериментально изучены лишь для небольшого числа вирусов. Известно, что ДНК представителей порядка Caudovirales (которые, кстати, имеют самые большие геномы среди архейных вирусов) кодирует часть или даже все компоненты аппарата репликации ДНК: ДНК-полимеразы, белки скользящего зажима ( PCNA ), праймазы и хеликазы. Имеющие более скромные размеры геномов вирусы архей, как правило, кодируют белки, которые необходимы для привлечения аппарата репликации клетки-хозяина. Стоит, однако, отметить, что в геномах многих вирусов, специфичных для архей, не удалось найти белки, связанные с репликацией ДНК, так что они либо полностью зависимы от аппарата репликации клетки-хозяина, либо используют уникальные, пока еще не изученные механизмы репликации ДНК. Например, удалось показать, что и инициация, и терминация репликации генома липотриксвируса AFV1 связаны с рекомбинационными процессами [1] .
Механизмы упаковки генома в капсид у вирусов архей детально не изучены. Тем не менее известно, что члены порядка Caudovirales имеют гомологи терминазы , которая упаковывает геномную ДНК в пустой капсид. Механизм формирования вирионов с использованием терминазы используют также вирусы бактерий и эукариот. Можно предположить, что упаковка генома в капсид у вирусов архей протекает так же, как и у бактериофагов и вирусов эукариот, а вот в плане репликации ДНК вирусы архей или целиком зависят от клетки-хозяина, либо используют уникальные, пока еще не изученные механизмы [1] .
Взаимодействие с клеткой-хозяином
Выход зрелых вирионов из клетки архей во многих случаях напоминает отпочковывание от эукариотической клетки-хозяина у вируса гриппа , ВИЧ и вируса Эбола . Когда вирион покидает клетку археи, он забирает с собой фрагмент ее мембраны, который становится дополнительной оболочкой поверх капсида. У некоторых вирусов архей финальные стадии созревания вирионов происходят уже после выхода из клетки, когда капсид претерпевает морфологические перестройки [1] .
У некоторых вирусов архей (а именно, представителей семейств Rudiviridae и Turriviridae ) все стадии созревания вириона проходят в цитоплазме клетки. Новые вирусные частицы покидают клетку через специальные структуры с семиосевой симметрией на их поверхности, которые получили название вирусассоциированных пирамид ( virus-associated pyramids, VAP ). VAP образуются на внутренней поверхности мембраны зараженной клетки, проходят сквозь ее поверхностный S-слой и открываются на финальных этапах инфекции, давая возможность вирионам выйти из клетки (рис. 4) [1] .
Выход некоторых вирусов архей из клетки сопровождается ее лизисом. К их числу относятся вирусы семейства Tristromaviridae , которые, хотя и развиваются целиком в цитоплазме, каким-то образом ухитряются приобрести липидную оболочку. Примечательно, что сифовирус ψM2 кодирует фермент псевдомуреинэндоизопептидазу , который разрушает псевдомуреиновую клеточную стенку археи [1] .
Эволюция и родственные связи
Вирусы, специфичные для архей, как правило, заражают только представителей типа Crenarchaeota . Они отличаются от всех других вирусов не только нестандартной морфологией вирионов, но и генетически: около 90% их генов не имеет гомологов в существующих базах данных. В геномах некоторых архейных вирусов не удалось найти ни одного белка, для которого существовал бы функционально охарактеризованный белок-гомолог.
Иногда, если гомологию не удается установить по нуклеотидным или аминокислотным последовательностям, на помощь приходят пространственные структуры. Действительно, для различных белков вирусов архей получили пространственные структуры, но ситуация от этого не стала яснее: оказалось, что во многих из них содержатся совершенно уникальные структурные мотивы. Более того, функции многих генов вирусов архей совершенно непонятны: так, оказалось, что фузелловирус SSV1 может спокойно обойтись без половины своих генов. Можно предположить, что такие неохарактеризованные гены-сироты кодируют белки, участвующие во взаимодействии вируса с клеткой архей, например, противодействующие системам CRISPR-Cas [1] .
Впрочем, многие вирусы архей родственны некоторым бактериофагам и вирусам эукариот. Однако вирусы, специфичные для архей, стоят особняком среди всех ДНК-содержащих вирусов. Более того, различные группы вирусов, специфичных для архей, неродственны друг другу и эволюционируют независимо друг от друга. Высказывается предположение, что некоторые группы специфичных для архей вирусов появились на заре эволюции клеточной жизни и были впоследствии утрачены бактериями и эукариотами. Другие группы специфичных архейных вирусов могли появиться в момент обособления домена архей или даже позже, в отдельных группах архей [1] .
Любопытно, что некоторые вирусы архей родственны лишенным капсидов мобильным генетическим элементам (например, плазмидам) . Эти вирусы, так же как и мобильные генетические элементы, имеют родственные гены основных белков репликативного аппарата [1] , [2] .
Несмотря на то, что мы только-только начинаем приподнимать завесу тайны над вирусами архей, многие уже известные особенности их морфологии или генетического аппарата поражают воображение. Несомненно, дальнейшее изучение вирусов архей при помощи метагеномики и других подходов подарит нам множество сюрпризов.
Литература
1. David Prangishvili, Dennis H. Bamford, Patrick Forterre, Jaime Iranzo, Eugene V. Koonin, Mart Krupovic. (2017). The enigmatic archaeal virosphere . Nat Rev Micro . 15 , 724-739;
2. Mart Krupovic, Virginija Cvirkaite-Krupovic, Jaime Iranzo, David Prangishvili, Eugene V. Koonin. (2018). Viruses of archaea: Structural, functional, environmental and evolutionary genomics . Virus Research . 244 , 181-193;
3. Maija K. Pietilä, Tatiana A. Demina, Nina S. Atanasova, Hanna M. Oksanen, Dennis H. Bamford. (2014). Archaeal viruses and bacteriophages: comparisons and contrasts . Trends in Microbiology . 22 , 334-344;
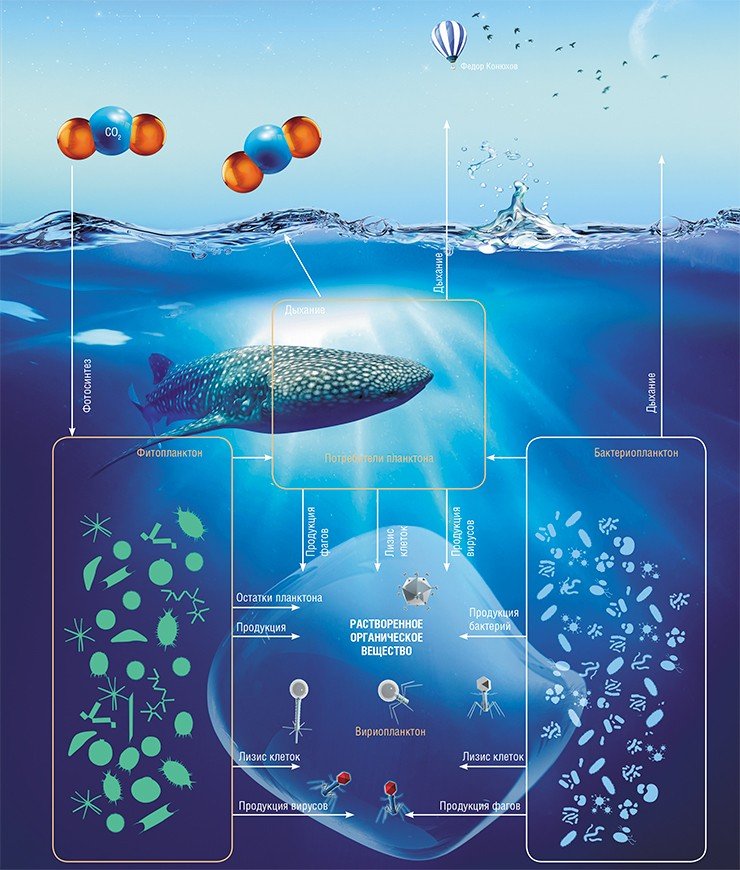
Везде, где есть жизнь, есть вирусы. Водная среда, занимающая бóльшую часть нашей планеты, в которой сосуществует огромное число различных организмов, создает прекрасные условия и для жизни вирусов. В водных экосистемах вирусы атакуют все живые организмы – от бактерий до китов. Оставим за рамками рассмотрения вирусы крупных организмов и останемся в микромире – в мире микроскопических организмов, которые являются основой пищевых цепей и, как выясняется, многих глобальных процессов
Первая информация о количестве вирусных частиц в водных экосистемах, потрясшая исследователей, была получена в 1989 г. (Bergh et al., 1989). Материал из проб морской воды был осажден центрифугированием прямо на сеточки с пленкой-подложкой и исследован в трансмиссионном (просвечивающем) электронном микроскопе. В одном миллилитре оказалось до 2,5 × 10 8 вирусных частиц, представленных в основном фагами с характерной морфологией (капсид-отросток, или голова-хвост), что в 10 3 —10 7 раз превышало концентрацию фагов, определенную путем традиционного высева проб на бактериальный газон (метод бляшек). Разница на порядки объясняется тем, что не все бактерии культивируются, и не все вирусы-фаги инфицируют именно бактерии.
![]()
Бактериофаги в глобальном круговороте
Во-первых, удаление из экосистемы части бактерий, уничтоженной фагами, уменьшает интенсивность перевода нерастворимого (взвешенного в водной толще и осаждающегося на дно) биогенного вещества (различных частиц, отмерших организмов и др.) – в растворенное (расщепленное, гидролизованное). Таким образом фаги нарушают классическую пищевую цепь. Этот процесс был назван вирусным шунтом (Wilheln, Suttle 1999). По оценкам авторов, через этот шунт может проходить до четверти первичной продукции углерода океана.
Количественная мультитрофическая модель, созданная авторским коллективом океанологов и математиков описывает влияние морских вирусов на микробиальные пищевые сети и процессы, проходящие в экосистемах. Согласно этой модели водные экосистемы, содержащие вирусы, будут иметь усиленный круговорот органического вещества, уменьшенный перенос этого вещества на более высокие трофические уровни и увеличенную валовую первичную продуктивность (Weitz et al., 2014). Авторы модели считают, что в оценках круговорота углерода и азота необходимо учитывать роль вирусов, так как они являются важной составляющей пищевых сетей и регулируют глобальные биогеохимические циклы.
Цианофаги – особый случай?
Цианобактерии (синезеленые водоросли), хотя и относятся к домену Bacteria, благодаря способности к фотосинтезу играют иную нежели гетеротрофные бактерии роль в водных экосистемах. Это одни из самых древних организмов. Они доминировали на ранних стадиях эволюции биосферы Земли и определяли биогеохимические циклы. Их бурное развитие вызвало изменение атмосферы, обогатило ее кислородом, что сделало возможным появление других организмов и направило эволюцию биосферы нашей планеты. Можно предположить, что цианофагов тогда еще не было.
Вирус морской диатомеи Chaetoceros debilis CdebDNAV – сохраняет инфекционность при широком диапазоне температур (от 20°С до -196°С) без добавления криопротекторов (Nagasaki, 2008).
Вирусы токсичной красной водоросли Heterosigma akashiwo сохраняют литическую активность в донных отложениях (Lawrence, 2002).
Цианофаги могут сохраняться в осадках до 100 лет (Suttle, 2000).
Гигантский вирус амеб Pithovirus sibericum – выделен из вечной мерзлоты возрастом 30 тыс. лет (Legendre et al., 2014)
И сегодня среди цианобактерий есть экстремофилы – виды, прекрасно существующие в горячих источниках, жарких пустынях, а также арктических и антарктических условиях. Если у таких видов есть цианофаги, то насколько они, эти цианофаги, уникальны?
![]()
Поскольку пресноводные нитчатые цианобактерии легко культивируются, именно из них и были выделены первые цианофаги – вирусные частицы в форме икосаэдра без хвоста диаметром 66 нм, о чем последовало короткое сообщение в Science (Safferman, Morris, 1963). Ими были инфицированы нитчатые цианобактерии Lyngbya, Plectonema и Phormidium. За последующие десять лет были выявлены фаги у других цианобактерий, в том числе пикопланктоных (Synechococcus, Microcystis) и нитчатых, формирующих гетероцисты (Anabena, Nostoc). Пробы для исследования были получены, в основном, из сточных вод и очистных сооружений.
За разнообразием – на Байкал!
Вирусы древнейших
Ферменты архей применяются в пищевой промышленности, так как могут работать при высоких температурах, а ДНК-полимераза археи Pyrococcus furiosus используется в ПЦР (полимеразной цепной реакции). Сами археи являются компонентом очистных сооружений, обеспечивая анаэробное разложение сточных вод; используются при обогащении руд ценных металлов. Ясно, что в промышленном производстве лизис архей вирусами – большая неприятность.
Структурные исследования вирусных капсидов показали, что бесхвостые икосаэдры, инфицирующие архей, бактерий и эукариот, имеют общего предка (Abrescia et al., 2012). Например, структурные сходства имеют белки оболочки нитчатых вирусов табачной мозаики, двух вирусов архей из рода Acidianus и вируса гепатита B. Несмотря на низкую гомологию аминокислотных последовательностей, белки вирусов архей могут иметь сходные элементы третичной структуры с вирусами других доменов (Dallas et al., 2014). А совсем недавно был описан нитевидный вирус гипертермофильной археи Pyrobaculum, который имеет уникальную среди ДНК-содержащих нитевидных вирусов структуру вириона. Его линейный геном заключен в трехслойный панцирь, состоящий из двух белковых слоев и дополнительной наружной оболочки. Вирион организован в виде суперспирали подобно вирусам Эбола и Марбург, но они являются РНК-содержащими (Rensen et al., 2016).
Каждую секунду в океане происходит 10 23 вирусных инфекций. Каждая инфекция имеет возможность для введения новой генетической информации в организм и в вирусное потомство, способствуя таким образом эволюции как сообществ хозяина, так и вирусов (Suttle, 2007). И хотя важность водных вирусов уже стала очевидной, на многие вопросы наука пока ответить не может.
Bergh Ø, Borsheim KY, Bratbak G, Heldal M. Abundance of viruses found in aquatic environments // Nature. 1989. V. 340. P. 467—468.
Hug L. A., Baker B. J., Anantharaman K. et al. A new view of the tree of life //Nature Microbiol. 2016. 11 Apr. N. 16048. DOI 110.1038.
Pietilä M. K., Demina T. A., Atanasova N. S., Oksanen H. M., Bamford D. H. Archaeal viruses and bacteriophages: comparisons and contrasts // Trends in Microbiology. 2014. V. 2. N. 6. P. 334—344.
Prangishvili D. The wonderful world of archaeal viruses // Annu. Rev. Microbiol. 2013. V. 67. P. 565–85.
Suttle C. A. Viruses in the sea // Nature. 2005. V. 437. P. 356—361.
Дрюккер В. В., Дутова Н. В. Бактериофаги как новое трофическое звено в экосистеме глубоководного озера Байкал // Докл. РАН. 2009. Т. 427, № 2. С. 277—281.
Автор благодарит Г. И. Филиппову (ЛИН СО РАН, Иркутск) за помощь в подготовке публикации
Читайте также:
 Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу.
Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу.
Copyright © Иммунитет и инфекции