По расположению жгутиков кишечная палочка
По существующей гипотезе, внутреннее, встроенное в цитоплазматическую мембрану кольцо базальной структуры - вращающийся ротор, окружено мембранными белками, имеющими определенным образом пространственно организованные отрицательные заряды - статор мотора. Протонодвижущая сила заставляет протоны проходить через базальную структуру внутрь клетки, при этом в какой-то момент они задерживаются в определенных участках ротора, придавая им положительный заряд, затем протоны уходят внутрь клетки. Заряженные участки расположены таким образом, что возникает сила притяжения между заряженными участками ротора и статора, кольцо начинает вращаться. установлено, что для полного оборота кольца через базальную структуру должно пройти 500-1000 протонов. Кольцо совершает около 300 оборотов в секунду. Нить жгутика закручена против ч.с. Если кольцо вращается против ч.с. нить ввинчивается в окружающий раствор. На работу жгутика бактерия тратит около 0,1% расходуемой энергии. У кишечной палочки 6-7 жгутиков. Бактерии не воспринимают направление силы тяжести из-за вращения клетки.
Важной гидродинамической характеристикой плывущего предмета является число Рейнольдса представляющее собой отношения сил инерции
к силам вязкости. С уменьшением размеров объекта это число уменьшается, поэтому водная среда является для бактерий вязкой, каковой для нас была бы патока. Подсчитанно, что плыть целенаправленно в одном направлении бактерия может не более 3 с, затем удары окружающих молекул разворачивают ее в случайном направлении.
Когда нить жгутика вращается против ч.с. (рис.2) бактерия движется по прямой - пробег. Скорость движения во время пробега для разных бактерий 20-80 мкм/с. Для кишечной палочки 30 мкм/с т.е. 10 см/ч. Пробег длится около секунды, после чего происходит переключение жгутикового мотора, который начинает вращаться по ходу ч.с. что приводит к остановке и развороту бактерии в случайном направлении. Переключение занимает менее 1 мс. У кишечной палочки при этом единая жгутиковая спираль распадается и бактерия совершает кувырок. Мотор вращается по ходу ч.с. около 0.1 с после чего снова переходит к вращению против ч.с.
Кишечная палочка имеет около 20 различных типов рецепторов, всего клетка кишечной палочки содержит около 25 тыс. молекул рецепторов. Рецепторы сконцентрированы на одном из полюсов.
У кишечной палочки в качестве важных компонентов сенсорной системы функционируют четыре так называемых метилакцепторных белка, которые являются рецепторами или получают сигналы от других рецепторов . Эти белки способны присоединять к заряженным глютаминным остаткам метильные группы или отдавать их за счет активности соответствующих ферментов, причем уровень метилирования белка соответствует концентрации эффектора. Не во всех таксисах так происходит.
Бактерии способны воспринимать изменение концентрации эффектора во времени, поскольку, если концентрация изменяется, она уже не соответствует уровню метилирования метилакцепторного белка. В этом случае сигнал поступает на определенные цитоплазматические белки, изменяя уровень их фосфорилирования, а эти белки передают сигнал уже на жгутиковый мотор, в результате чего изменяется длина пробега.
При увеличении концентрации аттрактанта длительность пробега возрастает ( а частота кувырков уменьшается), при увеличении концентрации реппелента длительность пробега уменьшается. При снижении концентрации эффекторов наблюдается обратный эффект. Это означает, что бактерии имеют элементарную память.
Обычно через 2-3 мин в результате процесса адаптации уровень метилирования белка приходит в соответствие с новой концентрацией эффектора и длина пробега возвращается к норме. Однако, если в среде создается градиент концентрации эффектора, в результате многочисленных изменений длины
пробега в зависимости от направления движения клетки бактерии постепенно приближаются к источнику аттрактанта или удаляются от реппелента (рис. 3)
Магнетотаксис - способность двигаться вдоль линий магнитного поля. В клетках таких бактерий находятся кристаллики железосодержащих минералов (напр. магнетит). Железо составляет около 3% сухой массы магнетобактерий. Это водные бактерии обитающие в пресных водоемах и море. В северном полушарии они плывут к северному полюсу и наоборот. Бактерии углубляются в воду т.к. магнетосомы ориентируются по результирующей вертикальной и горизонтальной составляющим магнитного поля. Чем ближе север тем круче они уходят в глубину в ил, где много пищи и мало кислорода.
Одним из крупнейших достижений человека на заре его развития было создание постоянного источника продуктов питания путем одомашнивания диких животных и возделывания растений. Создание разнообразных пород животных и сортов растений стало возможным благодаря разработке принципов искусственного отбора.
Окультуривание диких растений происходило независимо в различных географических областях и основывалась на местной флоре. Распространение культурных растений за пределы центра происхождения происходило как естественным путем, так и вследствие миграций населения. Так возникли первичные и вторичные центры происхождения и разнообразия культурных растений. Открытие центров происхождения культурных растений принадлежит выдающемуся русскому ученому Н.В.Вавилову, собравшему огромный семенной материал для использования в селекционной работе.
Было выделено восемь центров происхождения культурных растений:
1. Средиземноморский (спаржа, маслины. капуста, лук, клевер, мак, свекла, морковь)
2. Переднеазитский (инжир, миндаль, виноград, гранат, люцерна, рожь, дыня, роза)
3. Среднеазиатский (нут, абрикос, горох, груша, чечевица, лен, чеснок, мягкая пшеница)
4. Индо-Малайский (цитрусовые, хлебное дерево, огурец, манго, черный перец, кокосовая пальма, банан, баклажан)
5. Китайский (просо, редька, вишня, яблоко, гречиха, слива, соя, хурма)
6. Центрально-американский (тыква, фасоль, какао, авокадо, махорка, кукуруза, батат, хлопчатник)
7. Южноамериканский (табак, ананас, томат, картофель)
8. Абиссинский центр (банан, кофе, сорго, твердая пшеница)
В настоящее время выделяют 12 первичных центров происхождения культурных растений:
1. Китайско-Японский
2. Индонезийско-Индокитайский
3. Австралийский
4. Индостанский
5. Среднеазиатский
6. Переднеазиатский
7. Средиземноморский
8. Африканский
9. Европейско-Сибирский
10. Центральноамериканский
11. Южно-Американский
12. Северо-Американский
Циклы могут быть эндогенной природы и вызванные внешними периодическими изменениями
Циркадные ритмы привышают время генерации клетки или особи, т.е. лунные ритмы не встречаются у организмов период генерации которых меньше месяца, циркадные колебатели известны для тех клеток, время генерации которых превышает 24 часа. Свет, температура, длина светового дня - колебатели циклов.
Время генерации эукариотических клеток близко к 24 часам. В клетках Acetobularia лишенных ядра циркадианные ритмы фотосинтеза сохранялись 30 суток - цитоплазмы достаточно для поддержания активности колебателя.
Медоносные пчелы, найдя источник пищи, напр. в 3 ч дня, на следующий день прилетают к тому же самому месту точнов то же самое время - память времени. Если крыс, которые обычно едят в начале ночи, кормить только утром, то подъем активности ферментов в тонком кишечнике смещается на раннее утро и сохраняется в течении нескольких циклов после предоставления животному свободного доступа к пище. Благодаря изменении
длины дня защитные стратегии (зимняя спячка, эстивация, покой, диапауза) у животных включаются раньше наступления неблагоприятного времени. Изменяемое время регистрируется вплоть до минут и механизм такого измерения не выяснен.
У млекопитающих колебатель ритма расположен около зрительной хиазми, где к нему пододят нервные волокна, идущие от сетчатки.
У тараканов циркадианные осцилляторы находятся в левой и правой зрительной доле. У воробъя - эпифиз. У бабочек - сатурний циркадианный римт выхода имаго исчезает после обезглавливания куколок, но восстанавливается, если пересадить куколкам мозго в гемоцель брюшка. У Apliysia оба глаза являются автономными колебателями. У животных и человека имеется несколько осцилляторов. Осцилляторы нервной природы.
У дневных птиц типичный суточный профиль активности включает один пик после рассвета и второй перед закатом (пение, крики, кормление, ухаживание, спаривание, защита территории). Утренний пик больше - в это время происходит строительство гнезда.
Социальная синхронизация ритмов - крики и пение птиц разных особей служит времязадателем.
ПРИЛИВЫ
полусуточные приливы 12,4 ч преобладают на побережьях Атлантического океана, Северного моря и в нек районах Индийского океана. Амплитуда суточных пиков одинакова и меняется в течении синодического месяца (от новолуния до новолуния Т=29,53 дня). Максимальные приливы в сизигии - луна, земля и солнце на одной прямой - новолуние, минимальные приливы в квадратуре.
Суточные приливы - 24 ч
циркадный ритм Дрозофилы требует взаимодействия по крайней мере двух генов period (per) и timeless (tim) Их транскрипты накапливаются днем и уменьшаются ночью. Эти молекулярные ритмы результат негативной обратной связи, когда Per и Tim белки взаимодействуют проникая в ядро и ингибируют транскрипцию своих собственных генов. Свет быстро разрушает tim, транскрипция возобновляется.
Сферические формы (кокки) - шаровидные бактерии размером 0,5 - 1,0 мкм; по взаимнму расположению клеток различают микрококии, диплококки, стрептококки, тетракокки, сарцины и стафилококки.
Микрококки (лат. малый) - отдельно расположенные клетки или в виде "пакетов".
Диплококки (лат. двойной) - располагаются парами, так как клетки после деленияне расходятся.
Стрептококки (от греч. streptos - цепочка) - клетки округлой или продолговатой формы, составляющие цепочку вследствие деления клеток в одной плоскости и сохранения связи между ними в месте деления.
Сарцины (от лат. sarcina - связка, тюк) - располагаются в виде пакетов из 8-и и более кокков, так как они образуются при делении клетки в трех взаимно перпендикулярных плоскостях.
Стафилококки (от. греч. staphyle - виноградная гроздь) - кокки расположенные в виде грозди винограда в результате деления в различных плоскостях.
Палочковидные бактерии различаются пао размерам, форме концов клетки и взаимному расположению клеток. Длина клеток варьирует от 1,0 до 8,0 , толщина от 0,5 до 2,0 мкм. Палочки могут быть правильной (кишечная палочка) и неправильной (коринебактерии) формы, в том числе ветвящиеся, например актиномицеты. Слегка изогнутые палочки называют вибрионами (холерный вибрион). Большинство палочковидных бактерий располагаются беспорядочно, так как после деления клетки расходятся.
Риккетсии - мелкие грамотрицательные палочковидные бактерии (0,3 - 2,0 мкм), облигатные внутриклеточные паразиты. Размножаются делением в цитоплазме, а некоторые - ядре инфицированных клеток. Обитают в организме членистоногих (вшей, блох, клещей), которые являются их хозяевами или переносчиками. Форма и размер риккетсий могут изменяться (клетки неправильной формы, нитевидные) в зависимости от условий роста. Структура риккетсии не отличается от таковой грамотрицательной бактерии.
Хламидии - относятся к облигатным внутриклеточным кокковым грамотрицательным (иногда грамвариабельным) бактериям. Вне клеток хламидии имеют сферическую форму (0,3 мкм), метаболически неактивны и называются элементарными тельцами. В клеточной стенке элементарных телец имеется главный белок наружной мембраны и белок, содержащий большое количество цистеина. Хламидии размножаются только в живых клетках, их рассматривают как энергетических паразитов.
Элементарные тельца попадают к эпителиальную клетку путем эндоцитоза с формированием внутриклеточной вакуоли. Внутри клетки они увеличиваются и превращаются в делящиеся ретикулярные тельца, образуя скопления в вакуолях (включения). Из ретикулярных телец образуются элементарные тельца, которые выходят из клеток путем экзоцитоза или лизиса клетки.
Микоплазмы - мелкие бактерии (0,15 - 1,0 мкм), окруженные цитоплазматической мембраной и не имеющие клеточной стенки. Из-за отсутствия клеточной стенки микоплазмы осмотически чувствительны. Имеют разнообразную форму: кокковидную, нитевидную, колбовидную. Эти формы видны при фазово-контрастной микроскопии чистых культур микоплазм. Патогенные микоплазмы вызывают хронические инфекции - микоплазмозы.
Актиномицеты - ветвящиеся, нитевидные или палочковидные грамположительные бактерии. Свое название (от греч. actis - луч, mykes - гриб) они получили всвязи с образованием в пораженных тканях друз - гранул из плотно переплетенных нитей в виде лучей, отходящих от центра и заканчивающихся колбовидными утолщениями. Актиномицеты могут делиться путем фрагментации мицелия на клетки, похожие на палочковидные и кокковидные бактерии. На воздушных гифах актиномицетов могут образовываться споры, служащие для размножения. Споры актиномицетов обычно нетермостойки.
Общую филогенетическую ветвь с актиномицетами образуют так называемые нокарднеподобные (нокардиоформные) актиномицеты — собирательная группа палочковидных, неправильной формы бактерий. Их отдельные представители образуют ветвящиеся формы. К ним относят бактерии родов Corynebacterium, bdycobacterium, Hocardia и др.
Нокардиоподобные актиномицеты отличаются наличием в клеточной стенке Сахаров арабинозы, галактозы, а также миколовых кислот и больших количеств жирных кислот. Миколовые кислоты и липиды клеточных стенок обусловливают кислотоустойчивость бактерий, в частности, микобактерий туберкулеза и лепры (при окраске по Цилю-Нельсену они имеют красный цвет, а некислотоустойчивые бактерии и элементы ткани, мокроты - синий цвет).
Извитые формы - спиралевидные бактерии, например спириллы, имеющие вид штопорообразно извитых клеток. К патогенным спириллам относится возбудитель содоку (болезнь укуса крыс). К извитым также относятся кампилобактеры, хеликобактеры, имеющие изгибы как у крыла летящей чайки; близки к ним и такие бактерии, как спирохеты.
Спирохеты — тонкие, длинные, извитые (спиралевидной формы) бактерии, отличающиеся от спирилл подвижностью, обусловленной сгибательными изменениями клеток. Спирохеты имеют наружную мембрану клеточной стенки, окружающую протоплазматический цилиндр с цитоплазматической мембраной. Под наружной мембраной клеточной стенки (в периплазме) расположены периплазматические фибриллы (жгутики), которые как бы закручиваясь вокруг протоплазматического цилиндра спирохеты, придают ей винтообразную форму (первичные завитки спирохет). Фибриллы прикреплены к концам клетки и направлены навстречу друг другу. Другой конец фибрилл свободен. Число и расположение фибрилл варьируют у разных видов. Фибриллы участвуют в передвижении спирохет, придавая клеткам вращательное, сгибательное и поступательное движение. При этом спирохеты образуют петли, завитки, изгибы, которые названы вторичными завитками.
Спирохеты плохо воспринимают красители. Их окрашивают по методу Романовского—Гимзы или серебрением, а в живом виде исследуют с помощью разово-контрастнои или темнопольнои микроскопии.
Лептоспиры (род Leptospira) имеют завитки неглубокие и частые — в виде закрученной веревки. Концы этих спирохет изогнуты наподобие крючков с утолщениями на концах. Образуя вторичные завитки, они приобретают вид букв S или С; имеют 2 осевые нити. Патогенный представитель L. interrogates вызывает лептоспироз.
Жгутики бактерий определяют подвижность бактериальной клетки. Жгутики представляют собой тонкие нити, берущие начало от цитоплазматической мембраны, имеют большую длину, чем сама клетка. Толщина жгутиков 12-20 нм, длина 3-15 мкм. Они состоят из 3 частей: спиралевидной нити, крюка и базального тельца, содержащего стержень со специальными дисками (1 пара дисков - у грамположительных и 2 пары дисков - у грамотрицательных бактерий). Дисками жгутики прикреплены к цитоплазматической мембране и клеточной стенке. При этом создается эффект электромотора со стержнем-мотором, вращающим жгутик. Жгутики состоят из белка - флагеллина (от flagellum - жгутик); является Н-антигеном. Субъединицы флагеллина закручены в виде спирали. Число жгутиков у бактерий различных видов варьирует от одного (монотрих) у холерного вибриона до десятка и сотен жгутиков, отходящих по периметру бактерии (перитрих) у кишечной палочки, протея и др. Лофотрихи имеют пучок жгутиков на одном из концов клетки. Амфитрихи имеют по одному жгутику или пучку жгутиков на противоположных концах клетки.
Пили (фимбрии, ворсинки) - нитевидные образования, более тонкие и короткие (3-10нм х 0, 3-10мкм) , чем жгутики. Пили отходят от поверхности клетки и состоят из белка пилина, обладающего антигенной активностью. Различают пили, ответственные за адгезию, то есть за прикрепление бактерий к поражаемой клетке, а также пили, ответственные за питание, водносолевой обмен и половые (F-пили), или конъюгационные пили. Пили многочисленны - несколько сотен на клетку. Однако, половых пилей обычно бывает 1-3 на клетку: они образуются так называемыми "мужскими" клетками-донорами, содержащими трансмиссивные плазмиды (F-, R-, Col-плазмиды). Отличительной особенностью половых пилей является взаимодействие с особыми "мужскими" сферическими бактериофагами, которые интенсивно адсорбируются на половых пилях.
Споры - своебразная форма покоящихся фирмикутных бактерий, т.е. бактерий с грамположительным типом строения клеточной стенки. Споры образуются при неблагоприятных условиях существования бактерий (высушивание, дефицит питательных веществ и др.. Внутри бактериальной клетки образуется одна спора (эндоспора). Образование спор способствует сохранению вида и не является способом размножения, как у грибов. Спорообразующие бактерии рода Bacillus имеют споры, не превышающие диаметр клетки. Бактерии, у которых размер споры превышает диаметр клетки, называются клостридиями, например, бактерии рода Clostridium (лат. Clostridium - веретено). Споры кислотоустойчивы, поэтому окрашиваются по методу Ауески или по методу Циля-Нильсена в красный, а вегетативная клетка в синий цвет.
Форма спор может быть овальной, шаровидной; расположение в клетке -терминальное, т.е. на конце палочки (у возбудителя столбняка), субтерминальное - ближе к концу палочки (у возбудителей ботулиэма, газовой гангрены) и центральное (у сибиреязвенной бациллы). Спора долго сохраняется из-за наличия многослойной оболочки, дипиколината кальция, низкого содержания воды и вялых процессов метаболизмов. В благоприятных условиях споры прорастают, проходя три последовательные стадии: активация, инициация, прорастание.
8. Основные формы бактерий
Шаровидные бактерии (кокки) имеют обычно форму шара, но могут быть немного овальной или бобовидной формы. Кокки могут располагаться поодиночке (микрококки); попарно (диплококки); в виде цепочек (стрептококки) или виноградных гроздьев (стафилококки), пакетом (сарцины). Стрептококки могут вызывать ангину и рожистое воспаление, стафилококки — различные воспалительные и гнойные процессы.
Палочковидные бактерии самые распространенные. Палочки могут быть одиночными, соединяться попарно (диплобактерии) или в цепочки (стрептобактерии). К палочковидным относятся кишечная палочка, возбудители сальмонеллеза, дизентерии, брюшного тифа, туберкулеза и др. Некоторые палочковидные бактерии обладают способностью при неблагоприятных условиях образовывать споры. Спорообразующие палочки называют бациллами.Бациллы, напоминающие по форме веретено, называют клостридиями.
Спорообразование представляет собой сложный процесс. Споры существенно отличаются от обычной бактериальной клетки. Они имеют плотную оболочку и очень малое количество воды, им не требуются питательные вещества, а размножение полностью прекращается. Споры способны длительно выдерживать высушивание, высокие и низкие температуры и могут находиться в жизнеспособном состоянии десятки и сотни лет (споры сибирской язвы, ботулизма, столбняка и др.). Попав в благоприятную среду, споры прорастают, т. е. превращаются в обычную вегетативную размножающуюся форму.
Извитые бактерии могут быть в виде запятой — вибрионы, с несколькими завитками — спириллы, в виде тонкой извитой палочки — спирохеты. К вибрионам относится возбудитель холеры, а возбудитель сифилиса — спирохета.
9. Особенности морфологии риккетсий и хламидий
Риккетсии и хламидии относятся к классу Rickettsia облигат-ных внутриклеточных паразитов, который делится на два порядка: Rickettsiales и Chlamidiales.
Риккетсии представляют собой мелкие грамотрицательные микроорганизмы, характеризующиеся выраженным полиморфизмом—образуют кокковидные, палочковидные и нитевидные формы (рис. 22). Размеры риккетсии варьируют от 0,5 до 3-4 мкм, длина нитевидных форм достигает 10—40 мкм. Спор и капсул не образуют, окрашиваются по Здродовскому в красный цвет.
Хламидии имеют шаровидную, овоидную или палочковидную форму. Их размеры колеблются в пределах 0,2—1,5 мкм. Морфология и размеры хламидии зависят от стадии их внутриклеточного цикла развития, для которого характерно превращение небольшого шаровидного элементарного образования в крупное инициальное тельце с бинарным делением. Перед делением частицы хламидии обволакиваются образованием, напоминающим бактериальную капсулу. Хламидии окрашиваются"* по Романовскому—Гимзе, грамотрицательны, хорошо видны в прижизненных препаратах при фазово-контрастной микроскопии.
10. Строение и биология микоплазм.
К классу Mollicutes относится только один порядок Mycoplasmatales. Представители этого порядка — микоплазмы—
отличаются от бактерий отсутствием клеточной стенки. Вместо нее они содержат трехслойную липопротеидную цитоплазматическую мембрану. Размеры микоплазм колеблются в пределах 125—250 мкм. Они имеют форму круглых, овальных или нитевидных образований, грамотрицательны
Микоплазмы размножаются бинарным делением, подобно большинству бактерий, особенно после образования мелких кокковидных образований (элементарные тельца, ЭТ) в нитевидных структурах.
23 декабря 2014
- 9205
- 7,4
- 4
- 1
Несмотря на кажущуюся примитивность, бактерии не так уж просто устроены!
Бактерии — крохотные существа, и многие считают, что они очень просто устроены. Конечно, каждая бактерия — это всего лишь одна клетка, у которой нет отдельных частей тела, вроде ног или рук, нет глаз и носа, нет даже клеточного ядра. Но каким-то образом бактерии выживают и умудряются процветать с такими, казалось бы, ограниченными размерами и возможностями, да к тому же находить целое множество оригинальных решений для облегчения собственной жизни. Например, чтобы двигаться — то есть влиять на свое положение в пространстве самостоятельно, а не дожидаясь попутного течения, — бактерии придумывают самые удивительные ухищрения. Конечно, вы уже наверняка слышали о жгутиках бактерий. А что такое твитчинг? И как можно сдвинуть себя с места с помощью сахаров? Давайте присмотримся к бактериям чуть ближе. И сразу обратим внимание на то, из чего бактерии собирают себе средство передвижения.
Белковая диета
Среди подвижных бактерий больше всего тех, кто для движения использует белковые молекулы. Как они их применяют? Многие бактерии синтезируют специальные белки, из которых собирают подвижную ниточку — жгутик (рис. 1). Жгутик состоит из трех частей — филамента (собственно нити), крюка и базального тела. Каждая из этих частей сложена из белков. У хорошо изученной бактерии — кишечной палочки — белки, образующие филамент, называют флагеллинами и обозначают буквами Flg, Fli, Flh (от латинского слова flagellum — жгутик). Флагеллины складываются в нить, которая с помощью крюка крепится к базальному телу. Базальное тело — это что-то вроде якоря, который прочно закреплен в клеточной оболочке и может свободно вращаться по часовой стрелке или против. У бактерий может быть один или несколько жгутиков.
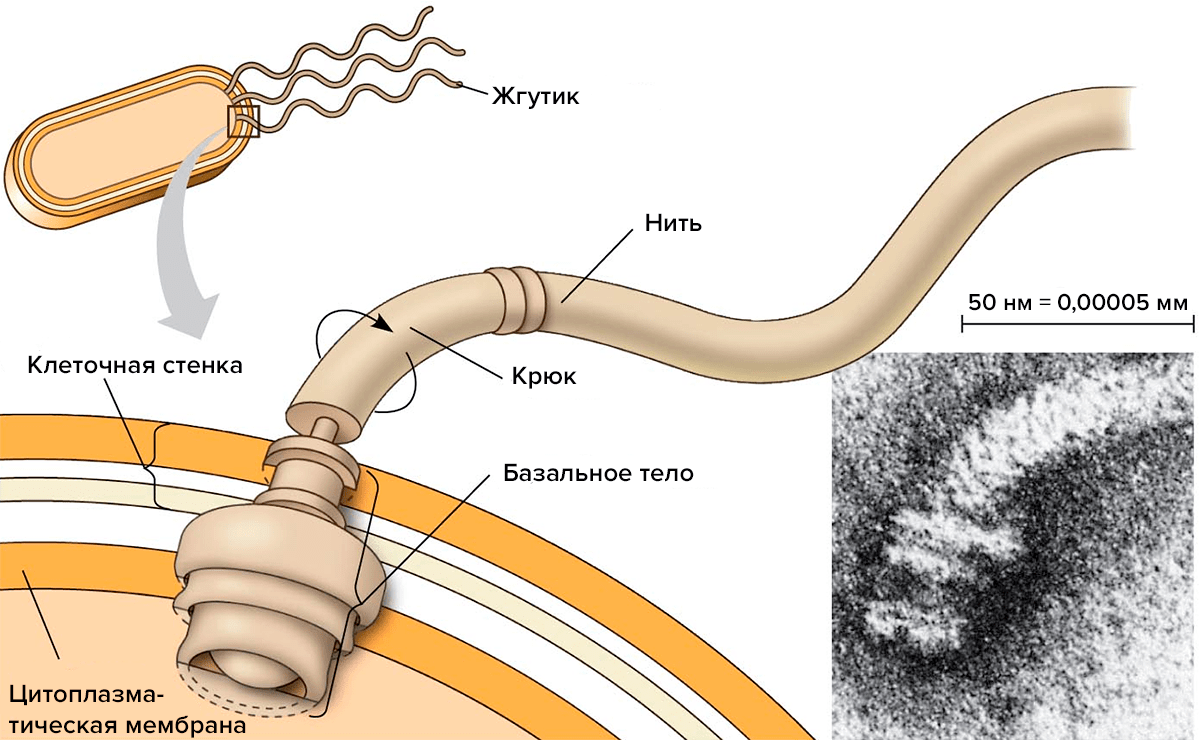
Рисунок 1. Строение жгутика.
Какие виды движения обеспечивает жгутик? Если бактерия находится в жидкой среде, то жгутик помогает ей плыть. Плавание — это самый быстрый способ передвижения. Причем, бактерия может неплохо управлять своим движением, меняя направление вращения базального тела: вращение базального тела по часовой стрелке толкает клетку в направлении от жгутика, а биения против часовой стрелки тянут клетку вслед за жгутиком.
А теперь представьте размахивание жгутиками на твердой поверхности, смоченной жидкостью. Бактерии будут не плыть, а расползаться в одной плоскости. Такое движение называется роением. Роение чаще бывает у бактерий, живущих в крупных колониях, — подвижные бактерии, находящиеся с краю, пытаются отодвинуться как можно дальше и основать свои собственные колонии.

Рисунок 2. Движение с помощью подтягивания к месту прикрепления пили.
Бактерии могут также создавать более короткие и просто устроенные нити, чем жгутики, — пили. Клетка может с помощью пили прикрепиться к чему-нибудь твердому, а потом подтянуться к месту крепления, разбирая эту нить, начиная от места крепления пили к клетке (рис. 2). Можно сказать, что клетка перемещается рывками. Подобный способ движения у одной клетки называется твитчинг (англ. twitch — дергать, тащить). А если так действует несколько скрепленных друг с другом бактерий, то они дружно скользят по твердой поверхности.
В оболочках клетки могут быть разнообразные белковые комплексы, например, обхватывающие клетку кольца из белков. Эти кольца крутятся, как гусеницы у гусеничного трактора, и помогают бактерии скользить по твердой поверхности. Такой способ подвижности есть у бактерии Flavobacterium johnsoniae.
У других бактерий есть белки, расположенные вдоль всей поверхности клетки. Эти белки создают продольные волны, и бактерия извивается и плывет в водной толще или скользит на твердом субстрате.
Очень многие микроорганизмы способны двигаться, но вот конкретный механизм или набор из нескольких механизмов у каждой специфичен. Поэтому, например, и говорят, что такое строение жгутика характерно для кишечной палочки, а у другой бактерии, тоже плывущей с помощью жгутика, все может быть устроено иначе — и ученые исследуют каждую бактерию по отдельности.
Как вы, наверное, заметили, пока что описанные способы движения позволяли бактерии плыть или скользить в зависимости от того, где она находится, — в жидкости или на твердой поверхности. Но скольжение может быть и единственным доступным способом передвижения.
Сахарный след
Многие бактерии выделяют наружу сахара. Смешиваясь с водой, сахара образуют слизь. Слизь облегчает движение клеток по твердой поверхности при использовании жгутиков.
Однако и сама слизь может быть источником движения. Представьте себе, что вы надуваете воздушный шарик. Внезапно шар вырывается из ваших рук и улетает под силой струи воздуха, резко выходящей из шарика. Подобным образом могут толкать себя и бактерии.
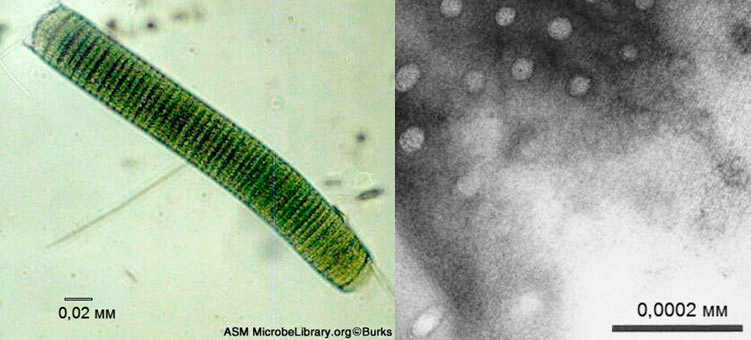
Рисунок 3. Oscillatoria princeps. Слева: Бактерии вида Oscillatoria princeps образуют нить. Справа: Большие поры на поверхности бактерии.
Выделение сахаров из специальных пор или биение жгутиков — это активные способы передвижения клетки. Бактерия взаимодействует с окружающей средой и активно отталкивается от воды или твердой поверхности. Но существует и пассивная подвижность, когда изменения внутри клетки приводят к ее перемещению благодаря внешним силам, например, току воды.
Газовые баллоны
Бактерии могут изменять свою плавучесть, накапливая внутри атмосферный воздух. Воздух все время диффундирует в толщу воды, а бактерии могут специально отбирать и накапливать молекулы разных газов в специальном баллоне, сложенном из белка. Таким образом клетка меняет свою плотность, начинает весить меньше и всплывает, выталкиваемая архимедовой силой. Если бактерии затем нужно погрузиться обратно, она может избавиться от воздуха или накопить внутри себя тяжелые сахара.
В какую сторону плыть?
Чтобы оказаться в самых подходящих для себя условиях, многие бактерии передвигаются не случайным образом, а целенаправленно, приближаясь к какому-нибудь приятному для себя объекту (например, еде или свету) или отплывая как можно дальше от неприятного (например, молекул, выделяемых другими бактериями). Такое целенаправленное движение называется таксисом. Чтобы распознавать сигналы из внешнего мира, бактерия синтезирует специальные белки — рецепторы, которые располагаются у нее на поверхности. Каждый вид рецепторов реагирует на свой стимул — молекулы еды, свет и так далее. Обнаружив свой стимул, рецептор передает сигнал о нем внутрь клетки.
Рисунок 4. Движение бактерии в направлении привлекающей ее пищи.
Если у клетки нет жгутика, то двигаться целенаправленно ей куда труднее. Но и тут можно что-то придумать. Например, газовые баллоны внутри клетки смещают бактерию вверх и вниз, то приближая ее к поверхности водоема, кислороду и свету, то погружая на дно.
Микроигра
Попробуйте расшифровать четыре слова, связанных с микробиологией, прослеживая движение бактерий к еде.
Каждая бактерия начинает двигаться от буквы, на которую указывает исходящая от бактерии стрелка. Затем бактерия меняет направление движения, согласно маленькой черной стрелочке у буквы. Если бактерия приближается к еде (красная точка), то она проплывает три клеточки (по горизонтали, вертикали или диагонали напрямую), снова поворачивает согласно направлению стрелки на этой клеточке и так далее. Если бактерия плывет в противоположную от еды сторону, она сдвигается только на одну клеточку. Если движение скорее нейтрально, то бактерия проплывает две клеточки.
| Патогенная кишечная палочка | |||||||
|---|---|---|---|---|---|---|---|
 | |||||||
| Научная классификация | |||||||
| Домен: | |||||||
| Учебный класс: | |||||||
| Бином имя | |||||||
| кишечная палочка Кишечной палочки ( / ˌ ɛ ʃ ə г ɪ K я ə K oʊ л ɪ / англизированный к / ˌ ɛ ʃ ə г ɪ K я ə K oʊ л aɪ / ; обычно сокращенно кишечной палочки ) является грамм - отрицательное , палочковидные бактерии , которая обычно находится в нижнем кишечнике на теплокровных организмов (эндотермических). Большинство кишечной палочки штаммов безвредны, но некоторые серотипы являются патогенными и могут привестисерьезным пищевым отравлением в организме человека , и иногда ответственность за отзыва продукции . E.coli , также ответственны за большинство случаев инфекций мочевых путей . Безвредные штаммы являются частью нормальной флоры в кишечнике , и может принести пользу своим хозяевам, производя витамин K 2 , и предотвращая создание патогенных бактерий в кишечнике. содержаниеВступлениеКишечная палочка и родственные бактерии составляют около 0,1% от кишечной флоры и фекально-оральная передачей является основным маршрутом , через который патогенные штаммы вызывают болезнь бактерии. Клетки способны выживать вне тела в течение ограниченного периода времени, что делает их идеальные индикаторные организмы для тестирования проб окружающей среды для фекального загрязнения . Бактерия также может быть легко и недорого , выращенной в лабораторных условиях, и интенсивно исследовалась на протяжении более 60 лет. E.coli , является наиболее широко изучен прокариотической моделью организма , а также важными виды в области биотехнологии и микробиологии , где он служил в качестве организма - хозяина для большинства работы с рекомбинантной ДНК . Немецкий врач - педиатр и бактериолог Эшерих обнаружили кишечную палочку в 1885 году, и теперь классифицируется как часть Enterobacteriaceae семейства гамма-протеобактерий . СеротипыПатогенные E.coli , штаммы могут быть классифицированы на основе элементов , которые могут вызывать иммунный ответ у животных, а именно:
Так , например, E.coli , штамм EDL933 имеет О157: H7 группы. Наружная мембрана из E.coli , клетки содержит миллионы липополисахарида (LPS) молекулы, которая состоит из:
Антиген вывода используются для серотипирования E.coli , и эти группы O обозначения идут от О1 до O181, за исключением некоторых групп , которые исторически были удалены, а именно O31, O47, O72, o67, O93 (ныне K84), O94, и O122; группы 174 до 181 являются предварительными (O174 = Ox3 и O175 = OX7) или находятся под следствием (176 до 181 являются STEC / VTEC). Кроме того , существуют подтипы для многих групп O ( например , O128ab и O128ac). Следует отметить , однако , что антитела по отношению к нескольким антигенам O перекрестной реакции с другими антигенами O и частично K антигенов не только из кишечной палочки , а также из других Escherichia видов и видов Enterobacteriaceae. Антиген O кодируется кластера генов РФБ. рол (CLD) ген кодирует регулятор длины липополисахарида О-цепи. Кислый капсульный полисахарид (КПС) представляет собой толстый, слизисто-подобные, слой полисахарида , который окружает некоторый патоген кишечной палочки . Есть две отдельных групп К-антиген группы, названных группы I и группы II (в то время как небольшие в промежутке между подмножеством (К3, К10, К54 и / K96) были классифицирован как группа III). Бывший (I), состоит из 100 кДа (крупных) капсульных полисахаридов, а вторые (II), связанные с внекишечными заболеваниями, находятся под 50 кДа в размере. Группа IK антигены можно найти только с определенным O-антигенами (О8, O9, O20 и O101 группа), они далее подразделены на основании отсутствия (IA, аналогично Klebsiella видов в структуре) или в присутствии (МБ) аминосахар и некоторые группы I K-антигены присоединены к липидному а-ядру липополисахарида (к LPS ), аналогичным образом на антигены O (и , будучи конструктивно идентичны антигенами O в некоторых случаях рассматриваются только как K антигены , когда экспрессировали совместно с другим аутентичным антигеном O). Группа II K антигены близко напоминают те , в грам-положительных бактерий и значительно отличаются по составу и далее подразделены в соответствии с их кислотных компонентов, как правило , 20-50% от цепей CPS связаны с фосфолипидами. В общей сложности есть 60 различных K антигены, которые были признаны (К1, K2A / переменного тока, К3, К4, К5, К6, К7 (= K56), К8, К9 (= O104), К10, К11, К12 (K82), К13 (= К20 и = K23), К14, К15, К16, K18a, K18ab (= К22), К19, К24, К26, K27, K28, K29, K30, K31, K34, K37, K39, K40, K41, K42 , K43, K44, K45, K46, K47, K49 (o46), К50, К51, К52, К53, К54 (= K96), К55, K74, K84, K85ab / переменного тока (= O141), K87 (= O32), K92, K93, K95, K97, K98, K100, K101, K102, K103, KX104, KX105, и KX106). Антиген Н является основным компонентом жгутиков, участвует в кишечной палочке движения. Это , как правило , кодируется FLIC гена. Есть 53 H определены антигены, пронумерованных от H1 до H56 (H13 и H22 не были E.coli , антигены , а из Citrobacter freundii и Н50 было установлено, что такой же , как Н10). Роль в болезниВ организме человека и домашних животных , вирулентные штаммы кишечной палочки могут вызывать различные заболевания. В организме человека: гастроэнтерит , инфекции мочевых путей , а также новорожденных менингит . В редких случаях, вирулентные штаммы также ответственны за гемолитической-уремический синдром , перитонит , мастит , сепсис и грамотрицательной пневмонии .
Некоторые штаммы кишечной палочки , такие как O157: H7 , O104: H4 , O121 , O26 , O103 , O111 , O145 и O104: H21 , производят потенциально летальные токсины . Пищевое отравление , вызванное кишечной палочки может привести от употребления в пищу немытые овощи или плохо разделывали и плохо приготовленное мясо. O157: H7 также печально известен за причинение серьезных и даже угрожающих жизни осложнений , таких как гемолитическая-уремический синдром . Этот штамм связан с США 2006 кишечной палочкой вспышки из - за свежий шпинат. O104: Н4 штамм одинаково вирулентный. Антибиотик и вспомогательные протоколы лечения для него не так хорошо развиты (она имеет возможность быть очень Энтерогеморрагическим как O157: H7, вызывая кровавый понос, но также и более энтероагрегативный, то есть он хорошо и комков придерживается кишечных мембран). Это напряжение за смертоносную июнь 2011 кишечной палочку вспышки в Европе. Степень тяжести заболевания значительно варьируется; это может привести к летальному исходу, особенно для маленьких детей, пожилых людей или ослабленного иммунитета, но чаще мягкие. Ранее, плохие гигиенические способы приготовления мяса в Шотландии погибли семь человек , в 1996 году из - за кишечной палочки отравления, и оставили сотни других зараженных. E.coli , могут питать оба термостабильных и термолабильных энтеротоксинов . Последние, называется LT, содержат одну субъединицу и пять B субъединиц , расположенную в один голотоксин, и очень близки по структуре и функции холерных токсинов. В B субъединиц помощь в соблюдении и вступления токсина в кишечных клеток - хозяев, в то время как субъединица расщепляется и предотвращает клетки поглощать воду, вызывая диарею . LT секретируется секреции пути 2 типа. Если E.coli , бактерия избежать кишечного тракта через перфорацию (например , из язвы , в разорванном приложении , или из - за хирургическую ошибку ) и введи живот, они обычно вызывают перитонит , который может быть смертельным без оперативного лечения. Тем не менее, кишечная палочка чрезвычайно чувствительна к таким антибиотикам , как стрептомицин или гентамицин . Последние исследования показывают , лечение энтеропатогенной кишечной палочки с антибиотиками не может улучшить исход заболевания, так как это может значительно увеличить вероятность развития гемолитического-уремического синдрома. Слизистая оболочка кишечника-ассоциированная кишечная палочка наблюдается в повышенных количествах в воспалительных заболеваниях кишечника , болезнь Кроны и язвенном колите . Инвазивные штаммы E.coli , существуют в больших количествах в воспаленной ткани, а количество бактерий в воспаленных регионах коррелирует с тяжестью воспаления кишечника. Желудочно-кишечные инфекции могут привести организм развивать память Т-клетки атаковать кишечник микробы, которые находятся в желудочно-кишечном тракте. Пищевое отравление может вызвать иммунный ответ на микробные бактерии кишечника. Некоторые исследователи предполагают, что это может привести к воспалительному заболеванию кишечника. Энтеросолюбильные кишечная палочка (ЭК) классифицируется на основе серологических характеристик и свойств вирулентности. Основные патотипы кишечной палочки , которые вызывают диарею, перечислены ниже.
| |||||||