
Паразитические черви процесс эволюции
Закончим эту главу рассмотрением вопроса о времени возникновения паразитизма в ходе эволюции жизни в условиях, способствующих его- возникновению. Вопрос о том, когда в ходе эволюции жизни на земле мог впервые возникнуть паразитизм, естественно, не поддается точному определению, но позволяет высказать по этому поводу некоторые косвенные соображения.
Пространственные и пищевые взаимоотношения (хозяин как место обитания и источник пищи паразита) в паре паразит — хозяин таковы, что паразиты обыкновенно значительно мельче хозяина. Линейные размеры паразита, как правило, не менее чем в десятки, а объемные, следовательно, в сотни раз меньше размеров хозяина. Отсюда напрашивается вывод, что паразитизм мог развиться в животном царстве в широком масштабе лишь после того, как животные организмы успели эволюционировать в значительной степени и дать целый ряд форм, существенно различающихся своими размерами. Только тогда меньшие формы получили возможность поселиться на больших или внутри их.
Несколько более конкретно можно подойти к вопросу о том, не когда появился паразитизм, а когда он впервые получал более ила менее широкое распространение. Прежде всего начали, вероятно, приспособляться к паразитизму простейшие, в чем им способствовали их микроскопические размеры. Однако и среди них паразитизм как более или менее широко распространенное явление мог развиться не ранее, чем появились различные группы свободно живущих червей. В самом деле, среди простейших отсутствуют формы, паразитирующие у губок, и лишь в ъвде редчайших исключений имеются простейшие, паразитирующие у кишечнополостных (жгутиконосцы Trypanophis в гастральной полости сифонофор, Amoeba hydroxena, инфузории Trichodina и Кегопа на гидре, инфузории Foettingeriidae в актиниях). Только начиная с червей, а в частности с кольчатых червей, мы наталкиваемся на широкое "приспособление простейших к паразитизму. В аннелидах и других высших червях имеются многочисленные инфузории, сотни видов грегарин, кокцидшь микроспоридии, Actinomyxidia, многие виды Peridinea (Haplozoon и др. в Polychaeta). Итак, время более широкого развития паразитизма среди одноклеточных мы можем относительно определить как совпадающее пли последующее за эволюцией кольчатых червей.
Развитие паразитизма у паразитических червей относится уже, вероятно, к той части палеонтологической летописи, которая нам известна, а именно: сопутствует или следует за появлением позвоночных. В самом деле, все представители современных паразитических гельминтов в половозрелом состоянии, как правило (кроме очень немногих сосальщиков и нематод), живут в позвоночных. В беспозвоночных встречаются только личиночные стадии их, для которых моллюски и членистоногие служат промежуточными хозяевами. Но самое явление развития с промежуточными хозяевами есть явление вторичное, которому предшествовало развитие простое, без смены хозяев. Таким образом, беспозвоночные включились в цикл гельминтов лишь позже, чем они стали паразитами позвоночных.
Путем этих рассуждений мы приходим к выводу, что паразитические черви появились не ранее силура—девона. Правда, среди нематод имеется несколько десятков видов, паразитирующих в перепончатокрылых, жуках и др., но развитие их должно быть отнесено к гораздо более поздним эпохам, к мезозою, так как сами хозяева их начинают появляться не ранее верхних горизонтов палеозоя. Большая группа паразитов (до 40 000 видов) образуется в среде двукрылых и перепончатокрылых (та- хины, наездники), но эволюцию этих паразитических форм можно отнести не ранее как к юрской эпохе, так как самые отряды двукрылых и перепончатокрылых ведут свое начало с юры. Возраст некоторых паразитических раков труднее поддается определению, так как среди них иногда целые группы (Bopyridae, Entoniscidae. многие Cirripedia) приспособились к жизни на или внутри других ракообразных или даже кольчатых червей (Xenocoeloma, Herpyllobius и др. из Copepoda), а также в иглокожих и в клоакальной полости асцидий.
В последнее время относительно отдельных видов паразитов накопились более конкретные данные, которые мы приводим, так как они не лишены интереса. Наиболее древние несомненные находки паразитов в ископаемом состоянии относятся к видоизмененным кольчатым червям, Myzostomidae, образующим галлы, или так называемые зооцецидии, на теле морских лилий. Такйе"галлы найдены не только у современных морских лилий, но и у ископаемых вплоть доj^cKoro периода.
В балтийском янтаре, т. е. в олигоцене, не раз были найдены насекомые с выходящими из их тела нематодами. В том же янтаре была обнаружена блоха Palaeopsylla clebsiana. Отсутствие у нее глаз, указывает на то, что она прошла уже долгий предварительный путь эволюции в паразитическом состоянии.
Павловский (1946 а) приводит целый ряд любопытных данных о наиболее давних сведениях относительно некоторых паразитов человека. Наиболее древней находкой является обнаружение объизвествленных яиц кровяной двуустки (Schistosoma haematobium) в. почках мумии XX династии фараонов Египта (1200—1090 до н. э.). О глистных заболеваниях упоминается в вавилонско-ассирийских клинописных таблицах.
Рисунки на фигурных глиняных сосудах древних перуанцев говорят о том, что еще за много столетий до открытия Америки европейцами, население этого материка страдало от песчаной блохи (Sarcopsylla penetrans) и от лейшманиозов (болезнь Ута). Новые доказательства древности человеческих паразитов дает исследование кишечника двух трупов, найденных в торфяниках бывшей Восточной Пруссии. Шидат (Szidat, 1944) сообщает, например, о нахождении в кишечнике обоих трупов яиц аскариды, власоглава, а также, может быть, и лентеца широкого. Между тем обе означенные находки датируются ранним ледниковым периодом.
Наконец, в вопросе о происхождении паразитизма немалую роль играют и морфологические, и физиологические, и экологические особенности животных, становящихся хозяевами паразитов. Паразитофауна любого животного в ее современном составе отнюдь не представляет чего-нибудь строго постоянного и устойчивого: она может меняться как в сторону уменьшения, так и в сторону увеличения ее разнообразия. Один и тот же хозяин в разных местах и при разных условиях может содержать в значительной мере разную паразитофауну, становится хозяином ряда новых паразитов.
Этот процесс становления организма хозяином паразитов представляет большой интерес и рассматривался Догелем (1927 6), а затем подвергся специальному анализу со стороны Павловского (1946 а). Целый ряд экспериментальных данных показывает, что многие организмы лимитируются в приобретении ими новых паразитов не только какими-нибудь эндогенными причинами (специфичность), но в значительной мере рядом внешних условий, устранение которых ведет к тому, что организм становится хозяином новых для него видов паразитов. Таким образом, помимо действительного, или фактического, набора паразитов определенного хозяина, можно еще говорить о его потенциальной пара- зитофауне.
Понятие о потенциальных паразитах имеет большой теоретический и практический интерес. Теоретически оно затрагивает вопрос о приобретении организмом свойственного ему ныне набора паразитов в процессе эволюции. Практически потенциальные паразиты организма являются угрозой, всегда нависающей над организмом их возможного хозяина.
Разбирая факторы, способствующие заражению хозяина определенным паразитом, Павловский относит к числу их следующие условия.
Во-первых, факторы, предрасполагающие организм к заражению его темл__илп иными паразитами. Такими являются, с одной стороны, строение и ф\нкции организма" при которых возможно проникновение паразита в организм. С другой стороны, сюда относится характер биохимизма опосредуемого паразитом организма, при котором возможно" не только переживание паразита в новом хозяине, но и дальнейшее прохождение в нем жизненного цикла.
Во-вторых необходимо Действие факторов, устанавливающих биоцс- нотические .связи между организмом п era потенциальным паразитом (через посредство промежуточных хозяев или через иные способы инвазии) .
Первую категорию факторов мы можем называть эндогенными, зависящими лишь от самого организма хозяина, вторую — экзогеп- н ы м и, или зависящими от окружающих условий.
Наконец, третьим моментом становления животного хозяином паразитов является наступление условий, при которых факторы внешней среды, действующие и на восприимчивого хозяина и на вирулентного паразита, в конечном счете осуществляют возможность приживания вирулентного паразита, проникшего в организм чувствительного хозяина (Павловский и Гнездилов, 1939).
Сочетание всех этих разнообразных условий и их воздействие очень хорошо иллюстрируется экспериментами Павловского, Гнездилова и Талызина (1936, 1939) над плероцеркоидами Diphyllobothrium latum. Они паразитируют во внутренних органах хищных рыб (окунь, щука, налим и др.). При поедании одной более крупной рыбой другой, бо>лее мелкой,, зараженной плероцеркоидами, проглоченные вместе с добычей плероцер- коиды проходят сквозь стенку желудка проглотившей хищной рыбы и поселяются в полости тела. В то же время в мирных рыбах, как, например, в карпе, плероцеркоиды никогда не обнаруживаются. Однако при искусственном введении живых плероцеркоидов в желудок карпа они пролезают через стенки желудка в полость тела и там приживаются.
Отсюда ясно, что в данном случае круг промежуточных хозяев, плероцеркоидов среди рыб определяется не устройством рта и кишечника определенных видов рыб (рот карпа способен проглотить плероцеркоида) и не биохимизмом разных видов рыб (ибо плероцеркоиды, введенные в кишечник карпа экспериментальным путем, удачно приживаются в этой рыбе). Причины незараженности карпа (и других мирных рыб) плероцеркоидами, очевидно, экзогенного характера и зависят от отсутствия пищевой связи между карпом и содержащими плероцеркоидов рыбами (щука, налим и др.)> т. е., вернее, односторонностью этой связи: хищные рыбы поедают карпа, но не поедаются им.
Столь же легко, как у карпа, приживаются экспериментально введен ные плероцеркоиды и у_миноги, которая нормально никогда их не содержит. Но причина этого отсутствия, по-видимому, иная, чем у карпа,, п зависит от анатомо-физиологических особенностей пищеварительного аппарата миноги. Миноги нападают на разных рыб, причем своим круглым ртом присасываются к телу рыбы и перетирают ее ткани множе: ством роговых зубов ротовой воронки. Такому перетиранию должны подвергаться и плероцеркоиды, если они имеются в поедаемой миногой рыбе.
Однако возможности заражения плероцеркоидами новых хозяев гораздо более широки и далеко выходят за пределы класса рыб. Аналогичные вышеприведенным успешные опыты введения плероцеркоидов были проведены и с травяной лягушкой, жерлянкой, разными видами ящериц., ужами, гадюкой и даже черепахой.
Таким образом, и здесь причиной отсутствия плероцеркоидов в перечисленных животных служит не- действительная невозможность этих животных заражаться плероцеркоидами, а отсутствие пищевых связей между ними и нормальными хозяевами плероцеркоидов. Исключение составляет только черепаха, у которой отсутствие заражений может зависть не только от отсутствия пищевых связей (экзогенная причина), но : от строения желудка взрослых черепах (эндогенная причина), а имен- v заражение черепах удается лишь у молодых, годовалых черепах, "згда как хрящеватая плотность стенок желудка взрослой черепахи метает заражению плероцеркоидами.
Кроме того, опыты с ящерицами выявили наличие еще одного фак
: ра, а именно: температурного. Дело в том, чго опыты заражения яще- нц плероцеркоидами удавались лишь при комнатной температуре, т. е. рн температуре, которая приблизительно отвечает таковой нормального естообитания плероцеркоидов (температура пресноводных водоемов в летнее время). При помещении же ящериц в термостат с температурой 37° С в опытах миграции плероцеркоидов не происходило, и они погибали.
Другой пример влияния температуры, хотя и в обратном направлении, дают опыты Матова (MatofJ, 1944) по заражению трихинами обычно не свойственных им хозяев, а именно: различных амфибий, т. е. холоднокровных позвоночных. Заражая амфибий (жаб, саламандр) мускульными трихинами при температурах 18—25° С, Матов не добился никаких ясных результатов. Однако при содержании подопытных животных в термостате при 31—37° С личинки трихин выходили в их кишечнике из капсул, развивались в самцов и самок, а у саламандр дело доходило даже до откладки самками личинок и до развития мускульных трихин. Правда, в последнем случае постановка опытов была искусственной, так как самки трихин вводились автором прямо в полость тела саламандры.
Все эти примеры показывают нам, что потенциально весьма многие организмы могли бы стать хозяевами для гораздо более многочисленного контингента паразитов, чем тот, которым они в действительности обладают, и, наоборот, многие паразиты могли бы иметь потенциально гораздо более широкий круг хозяев. Кроме того, разобранные случаи говорят, в какой сильной мере приобретение паразитом новых хозяев зависит от комбинации факторов разного порядка и частью эндогенного, частью экзогенного характера: строение и функции организма хозяина, биоценотические связи хозяина с проспективным паразитом, наконец, условия среды, способствующие или не препятствующие проникновению паразита в организм хозяина (температура и др.) и приживанию его в последнем.
— болезни человека, животных и растений, вызываемые паразитическими червями — гельминтами (глистами). Гельминтозы в зависимости от видовой принадлежности их возбудителей, подразделяются на следующие группы: трематодозы (возбудители.
Паразит разрывает оболочку икринки, попадает в воду и вскоре распадается на части по
Полипы становятся свободноживущими и первые 4—5 дней питаются за счет запасов желтка.
(кишечнополостных, червей, моллюсков и т. д.). Довольно часто попадаются. отдельные.
Первый вариант борьбы с простейшими паразитами. Полынь - источник горького вкуса. Рекомендую с помощью полыни производить очищение организма от простейших микроорганизмов: одноклеточных жгутиковых, гемолитического стафилококка, протеуса.
Аскариды — крупные раздельнополые черви (), паразит
Плоские черви — группа организмов, которая объединяет большое количество беспозвоночных, не имеющих полости тела. В современном виде группа явнопарафилетическая, но современное состояние науки не позволяет разработать удовлетворительную строго филогенетическую систему. Рис.1Свободноживущий плоский червь Среда обитания плоских червей – пресные и соленые водоемы, правда, отдельные виды занимают влажные наземные местообитанияе. К типу плоские черви относиться большое количество паразитов позвоночных и беспозвоночныхживотных. В настоящее время описано около 25 000 видов, в России — более 3000 видов. Плоские черви по сравнению с другими червями эволюционно низшие. В настоящее время выделяют семь классов плоских червей: 1. Ресничные черви, 2. Моногенеи, 3. Ленточные черви (Cestoda), 4. Цестодообразные (Cestodaria), 5. Гирокотилиды (Gyrocotyloidea), 6. Трематоды (Trematoda), 7. Аспидогастры (Aspidogastrea). Ресничные черви, Трематоды и Ленточные черви преобладают. Плоские черви – первые двусторонне-симметричные животные с уплощенным в спинно-брюшном направлении телом. Плоские черви имеют хорошо дифференцированнуюи четко оформленный задний конец. Кроме того, можно различить верхнюю и нижнюю поверхности. Для свободноживущих плоских червей характерно передвижение ползком или вплавь. Эта способность обеспечивается кожно-мускульным мешком и ресничками. Паразитирующие черви при передвижении могут пользоваться присосками (передвигаются по типу гусеницы-землемера). Ленточные черви используют перистальтику кожно-мускульного мешка. Снаружи тело покрыто однослойным эпителием, возможно, реснитчатым (ресничные черви).У сосальщиков,моногеней, цестодообразных и ленточных червейпокровы состоят изтак называемого тегумента, который в ряде групп имеет в своем строении микроворсинки или хитиновые крючки. Под эпителием располагается мускульный мешок, состоящий из нескольких слоев мышечных клеток: наружного и внутреннего. Специальные органы дыхания отсутствуют. Газообмен у большинства червей происходит через покровы тела. Во всех группах, кроме цестодообразных и ленточных червей, имеется глотка, ведущая в кишку или в пищеварительную паренхиму. Кишка слепо замкнута и сообщается с окружающей средой только через ротовое отверстие. Кишечник может быть прямой или разветвленный, все зависит от размеров особи. Глоткуможно найти на брюшной поверхности, нередко посередине или ближе к заднему концу тела. Нервная система состоит из нервных узлов в передней части тела, мозговых ганглиев и отходящих от них нервных столбов с комиссурами. Органы чувств, как правило, представлены отдельными кожными ресничками - отростками чувствительных нервных клеток. Некоторые свободноживущие представители типа в процессе приспособления к условиям обитания приобрели светочувствительные пигментные глазки - примитивные органы зрения и органы равновесия. Выделительная система состоит из протонефридиев — ветвящихся каналов, соединяющихся в один или два выделительных канала. В своем большинстве плоские червиявляются гермафродитами. Мужская и женская половая система имеет очень разнообразное строение. Семенники, яичникии их различная организация, другие части половой системы (например, мужские копулятивные органы) используются для определения систематического положения. Рассмотрим основные классы плоских червей, имеющие наибольшее значение в природе.
Исходя из всего выше описанного можно сделать вывод о том, что паразитические представители типа плоские червы очень похожи. Цестоды и сосальщики характеризуются вторичной редукцией нервной, органов чувств, выделительной, дыхательной системы, отсутствие пигментации покровов тела. Все представители обитают в желудочно-кишечном тракте, на личиночной стадии меняют среду обитания, одного или нескольких хозяев. Все это объясняется общностью происхождения. Большинство зоологов полагают, что формы, сходные с современной планарией, представляют собой общего предка, из которого 600 - 700 миллионов лет назад и развилось все многообразие истинно многоклеточных. Но паразитический образ жизни оказал различное влияние на эволюцию сосальщиков и цестод. Развитие пошло разными путями. Ленточные черви - плоские черви, на которых паразитический образ жизни сказался значительно сильнее, чем на сосальщиках и турбелляриях. Сосальщики имеют сильнее редуцированный организм, более приспособленный для паразитизма. Наличие отдельной половой системы в каждом членике, возможность отрыва членика без повреждения всего организма, неограниченный рост в течении всей жизни – все это приспособления к паразитическому образу жизни, направленные на воспроизводство большего числа потомков для сохранения вида. Жизненный цикл довольно простой, заключается в превращении онкосферы в финну, а финны в ленточную половозрелую стадию. Финна не имеет особых приспособлений, не способна к активному передвижению и размножению. Сосальщики пошли по другому пути. С их паразитическим образом жизни связан сложный жизненный цикл, включающий три личиночных стадии: мирацидий, редия, церкария. Все личинки активны, в состоянии двигаться на встречу промежуточному хозяину. На стадии редии даже возможно размножение. Все стадии имеют морфологические, физиологические и репродуктивные адаптации к паразитическому образу жизни, которыеспособствует увеличению численности потомства на протяжении жизненного цикла.
Галактионов К.В., Добровольский А.А.
В книге рассматриваются происхождение и эволюция жизненных циклов одной из центральных групп паразитических плоских червей — трематод. Приводится краткое описание особенностей строения и развития партеногенетических и гермафродитного поколений этих паразитов, дана характеристика основных тенденций их морфологической эволюции. На этой основе предпринят анализ многообразия конкретных путей реализации жизненных циклов трематод в конкретных биотопах. Проведена типологизация жизненных циклов с использованием модифицированной классификации В.А. Догеля и прослежена приуроченность различных типов жизненных циклов к разным типам экосистем. Дана характеристика структуры популяций трематод, которая рассматривается как производная фазовой организации их жизненных циклов, реализующихся в тех или иных эко- и биотопических системах. В заключительной главе обосновывается система взглядов авторов на эволюцию жизненных циклов трематод, выделяются основные филетические стволы и доказывается непродуктивность традиционных дихотомических и кладистических подходов к филогенетике паразитов со сложными жизненными циклами. Становление трематод в процессе эволюции рассматривается как результат сложных эволюционных преобразований их жизненных циклов, которые характеризуются разнонаправленностью путей адаптациогенеза отдельных фаз и поколений, их составляющих.
Пользователь обязуется не осуществлять (и не пытаться получить) доступ к каким-либо материалам ЭБ иным способом, кроме как через интерфейс Сайта.
Пользователь обязуется не воспроизводить, не дублировать, не копировать, не продавать, не осуществлять торговые операции и не перепродавать материалы ЭБ для каких-либо целей.
| № | Название | Автор | Рубрика | Номер гранта | Текст |
|---|---|---|---|---|---|
| 1 | 2-регулярные решения нелинейных задач. Теория и численные методы. | Измаилов А.Ф., Третьяков А.А. | математика, механика, информатика | 99-01-14129 |  |
| 2 | 9-й Баховский коллоквиум по азотфиксации, посвященный памяти члена-корреспондента РАН В.Л. Кретовича. Москва, 24-25 января 1995 г. Тезисы докладов | Евстигнеева З.Г. и др. | биология и медицинские науки | 00001209 | |
| 3 | И.С. Аксакова: история славянофильской газеты: Исследования. Материалы. Постатейная роспись | Вихрова Н.Н. и др. | филология и искусствоведение | 17-04-16086 | |
| 4 | страницы истории российского академического кавказоведения конца XIX - начала XX в.: кавказоведческое наследие А.С. и П.С. Уваровых | Алиева А.И. | филология и искусствоведение | 17-04-16008 | |
| 5 | Acrothoracica, сверлящие ракообразные | Колбасов Г.А. | биология и медицинские науки | 08-04-07038-Д | |
- Книги, изданные при поддержке РФФИ
- Вестник РФФИ, издание на русском языке
- Вестник РФФИ, издание на английском языке
- Вестник РФФИ. Гуманитарные и общественные науки
- Научно-популярные статьи и фотоматериалы
- Аннотированные отчеты по проектам РФФИ

Россия, 119334, Москва, Ленинский проспект, 32а, 20-21 этаж
Телефон для справок: +7 (499) 941-01-15

Дата публикации: 14.05.2018 2018-05-14
Статья просмотрена: 144 раза
Ключевые слова: окончательный хозяин, внутренняя среда организма, паразитический образ жизни, паразитический червь, трематоды.
Цель работы: изучить наиболее важные особенности строения паразитических червей класса Trematodes, позволяющие данным паразитам существовать в организме окончательного хозяина.
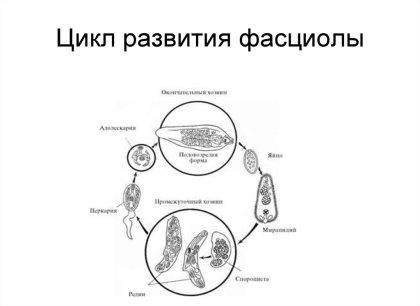
Взрослая половозрелая особь трематод имеет ряд специфических защитных механизмов и свойств, позволяющих данному классу вести паразитический образ жизни:
1) Трематоды, в зависимости от места локализации имеют разнообразную форму тела. Так трематоды вида Lyperosomum intermedium имеют удлиненное узкое тело, а трематоды вида Fasciola hepatica имеют уплощенное в дорсо-вентральном направлении листовидное вытянутое тело. Строение тела трематод обуславливает их локализацию в организме человека, позволяя избегать механического воздействия со стороны структур внутренних органов.
2) Характерной особенностью строения сосальщиков является наличие специальных органов прикрепления — 2 присосок, представляющих собой мускульные валики, образующиеся из кольцевых и радиальных мышечных волокон. За счет последовательных сокращений мускулатуры присосок осуществляется прикрепление паразитов к стенкам внутренних органов и сосудов. При этом в зависимости от расположения червей в организме изменяется степень развития присосок. Так наибольшего развития присоски достигают у тех трематод, которые обитают в кишке (Leucochloridium, Echinostomatida). Данный механизм позволяет червям защитить свой организм от выбрасывания во внешнюю среду.
3) Одной из важнейших структур трематод является покров тела, представленный тегументом. Тегумент подразделяется на наружный и внутренний. Наружный слой представлен безъядерным синцитием, а внутренний — участками цитоплазмы, несущими ядра. В составе тегумента содержатся белки, нейтральные и кислые мукосахариды. Они играют важную роль, как ингибиторы пищеварительных ферментов окончательного хозяина.
4) В ходе эволюции фасциолы утратили большинство органов чувств, лишь некоторые отдельные виды червей (Dicrocoelium) сохранили так называемые сенсиллы (простейшие органы чувств) на ротовой присоске. Иногда присутствуют рудименты глаз у церкариев (у Diplodiscus) или даже развитые глаза у паразита морских рыб (Neophasis oculatus). Отсутствие органов чувств у паразитических червей увеличивает приспособляемость организма к условиям внутренней среды хозяина.
5) Практически все сосальщики являются гермафродитами. Мужская половая система состоит из пары семенников, двух семяпроводов, сливающихся в семяизвергательный канал, и цирруса (копулятивного органа). Женская половая система представлена яичником, желточниками, семяприемником, который открывается в оотип (место, где осуществляется оплодотворение и происходит окончательное формирование яиц). Гермафродитизм у трематод является важным адаптационным механизмом, необходимым для существования внутри окончательного хозяина, так как у червей отсутствует необходимость в поиске полового партнера, а мощное развитие полового аппарата способствует увеличению количества новых особей.
Вывод: Таким образом, паразитические черви класса Trematodes имеют разнообразные адаптационные механизмы, необходимые для существования внутри организма окончательного хозяина. Данный класс паразитических червей представляет серьезную опасность для жизни и здоровья человека, так как может вызвать серьезные заболевания, такие как легочно-сердечная недостаточность, абсцессы головного мозга, менингоэнцефалит, абсцесс печени. Но знание жизненных циклов и морфологии данных паразитов способствуют разработке наиболее эффективных мер профилактики и защиты от опасных паразитических червей — трематод.
- Гинецинская Т. А. Трематоды, их жизненные циклы, биология и эволюция / АН СССР. — Л.: Наука, Ленингр. отд-ние, 1968. — 412,2)
- Слюсарев А. А., Жукова С. В. Биология. — Киев: Вища школа, 1987. — 412 с.
- Галактионов К. В., Добровольский А. А. Происхождение и эволюция жизненных циклов трематод / Рос. акад. наук, Зоол. ин-т, Кол. науч. центр РАН, Мурм. мор.-биол. ин-т. — СПб.: Наука, 1998. — 404, [16] с
- Морозова Д. А. Трематоды рыб озера Тана: Эфиопия: диссертация … кандидата биологических наук: 03.02.11 / Морозова Дарья Андреевна; (Место защиты: Ин-т проблем экологии и эволюции им. А. Н. Северцова РАН). — Борок, 2011. — 176 с.
- Скрябин К. И. Трематоды животных и человека. — М.: Академия наук, 1950. — 256 с
Список использованных источников
Наиболее распространенные виды гельминтов, паразитирующих в организме человека, принадлежат к классам сосальщиков ( Trematoda ), ленточных червей ( Cestoda ) и круглых червей ( Nematoda ). Систематика и биология паразитических червей человека и теплокровных животных являются одним из разработанных разделов паразитологии, что в значительной степени облегчает нашу задачу.
Класс сосальщиков объединяет несколько тысяч видов, при этом все они являются паразитами. По-видимому, это — древние паразиты, так как значительная часть их паразитирует в организме моллюсков, рыб и земноводных (эк- то- и эндопаразиты). Биологический цикл их имеет две модификации: одна часть трематод развивается прямым путем, без промежуточного хозяина и без партеногении; другая часть развивается с партеногенией в теле промежуточного хозяина (одного или двух). Сосальщики, паразитирующие в организме человека, относятся к отряду переднеротых ( Prosostomata ) и имеют сложный цикл развития. Половозрелые особи обитают в кишечниках теплокровных животных — птиц и млекопитающих; личинки проходят стадию развития в организме двух промежуточных хозяев, из которых первый — обычно моллюск, а второй — рыба или моллюск.
Эволюцию трематод можно представить следующим образом. По-видимому, наиболее древние виды являлись (и являются ныне) паразитами моллюсков и рыб. В ходе приспособления к паразитическому образу жизни у значительной части трематод сложился биологический цикл развития: рыбы — моллюски — рыбы. Поедание рыбами моллюсков обусловило и смену хозяев взрослой и личиночной стадии паразитов, что хорошо обеспечивало сохранение и распространение паразитического вида.
С появлением теплокровных — птиц и млекопитающих, которые питались рыбой и моллюсками, эволюция многих сосальщиков пошла по пути приспособления к организму новых хозяев. Этим и объясняется обилие паразитических сосальщиков у болотных и водоплавающих птиц, водяных крыс, бобров, тюленей, китообразных и подобных им животных. При этом цикл развития паразита изменился в одном из двух направлений:
а) либо место рыбы как окончательного хозяина занял соответствующий вид теплокровного животного,
б) либо рыба сохранилась как второй промежуточный хозяин. В первом случае заражение происходило через воду или при поедании моллюсков, во втором случае — при поедании рыбы, а иногда и другого водного животного (ракообразного, моллюска), которые являются вторым промежуточным хозяином. В зависимости от местных условий эволюция паразитов могла идти либо по пути строгой адаптации к определенным видам животных-хозяев, либо, что было гораздо чаще, по пути приспособления к более широкому кругу хозяев, без узкой специализации.
Первобытный человек, пока он вел кочевой образ жизни, мог лишь случайно заразиться трематодами — теми же видами, которые приспособились к широкому кругу хозяев (полигостальные паразиты). Заражение могло происходить при питье воды или употреблении в пищу сырой рыбы, моллюсков, ракообразных. Пока эти заболевания носили случайный характер, они не могли влиять на данный сложивший биоценоз. Примерами такого рода биоценозов, где человек является случайным звеном, могут служить гельминтозы типа гимастлоза (цикл: морские птицы — моллюски—морские птицы), описторхоза виверры (виверры — моллюски— рыбы — виверры), в первом случае человек заражается при поедании сырых моллюсков, во втором случае — при поедании рыбы.
Положение существенно изменилось, когда, люди перешли к оседлому образу жизни, поселились на берегах водоемов (рек, озер) и когда рыбная ловля стала одним из основных средств добычи пищи. В этих условиях некоторые виды гельминтов, особенно те из них, которые не были узко адаптированы к организму определенного вида животных, а являлись полигостальными паразитами, могли приспособиться к новому хозяину. Однако такими новыми хозяевами для них явились не человек, а домашние животные, притом плотоядные. Это обусловливалось следующим! причинами. На той стадии развития, о которой идет речь, человек был хорошо знаком с огнем и вряд ли часто употреблял в пищу сырую рыбу. Зато домашние плотоядные — кошки, собаки — в изобилии получали рыбу, главным образом ее внутренности. В силу этого весьма вероятной стала циркуляция паразита по типу: кошки (собаки) — моллюски — рыбы — кошки (собаки). Естественно, что эти благоприятные для паразита условия явились причиной образования новых видов глистов.
В настоящее время существует немалое число таких гельминтозов, которые отнесены в группу рыбных. Назовем некоторые из них: описторхоз, клонорхоз, нанофиетоз, стелланхазмоз, гетерофиоз. При всех этих гельминтозах главными хозяевами паразита являются кошки и собаки. Некоторые из рыбных трематодозов еще не стали болезнями только домашних плотоядных и сохранили более древние природные связи. Например, глист Pseudamphistomum truncatum паразитирует в печени тюленей и кошек; первые, по-видимому, прежде являлись единственными хозяевами. Некоторые паразиты приспособились к организму далеко для ловли рыбы. Иногда в эпизоотический процесс вовлекалось еще одно домашнее животное — свинья, которая всеядна и нередко питается рыбными отбросами. Примером подобного биоценоза может служить клонорхоз в Китае. Все эти гельминты могут вызывать заболевания человека, однако не человек, а домашние плотоядные животные являются основными резервуарами паразитов.
Из сказанного ясно, что ни один из гельминтов (рыбные трематоды) не смог стать чисто человеческой инвазией.
С переходом человека к оседлому образу жизни и заселению берегов водоемов стали возможными адаптация глистов к организму домашних животных и человека и образование таким образом новых видов трематод. По-видимому, таким путем возникли некоторые гельминтозы домашнего скота — фасциолезы, плагиорхоз. Циркуляция паразита осуществляется следующим путем: домашний скот — попадание фекалий в водоемы — пресноводные моллюски — выход инвазионных личинок в воду — проглатывание их домашним скотом при водопое. Эволюция многих видов пошла по пути развития личинки вне воды — в организме сухопутной улитки. Такой цикл был более налажен, так как фекалии животных попадали на почву (траву) и значительно реже — в воду. Примером подобного гельминтоза является дикроцелиоз. И фасциолезы, и дикроцелиоз являются гельминтозами домашних травоядных. Ни один из этих гельминтозов не стал чисто человеческой инвазией, хотя, казалось бы, для этого имелись все условия. Этот факт не совсем понятен, так как имеется не такое уж малое число инфекций человека, передающихся преимущественно или исключительно через почву. По-видимому, не всякая возможность реализуется в ходе эволюции, а может быть, данная возможность является только кажущейся вследствие недостаточности наших знаний.
Из числа трематод моногостальными паразитами человека стало только несколько видов шистосом, сохранивших промежуточного хозяина в виде моллюсков. Среди кровяных двуусток имеются паразиты диких птиц (например, трихобильгарциоз) и млекопитающих ( S . matthei , S . bovis , S . spindale ). Некоторые из шистосом, поражающих широкий круг млекопитающих, способны заражать и человека, например возбудитель японского шистосомоза — S . japonica , возможно, S . intercalatum (эндемик некоторых районов экваториальной Африки). Особенностью их цикла развития является активное внедрение личинок в кожные покровы главного отстоящих друг от друга животных, например глист Metagonimus yokogawai паразитирует в организме пеликанов, кошек и собак. Здесь можно высказать два предположения: либо пеликаны были первоначальными хозяевами, либо же стали таковыми позже, с тех пор как китайцы приручили их и стали использовать хозяина. По-видимому, развитие этих гельминтов пошло по двум путям: а) приспособление к организму домашних травоядных, которое сопровождалось переменой механизма — заражение теперь происходило алиментарным путем (водопой), так как внедрение их из воды в толстую кожу домашних травоядных было маловероятным; б) приспособление к организму человека с сохранением прежнего механизма заражения (внедрение в кожу при купании или работе в воде и в слизистые оболочки при питье воды); примерами подобных трематод являются S . mansoni (возбудитель кишечного шистосомоза), S . haematobium (возбудитель мочеполового шистосомоза).
В настоящее время шистосомозы по распространенности являются вторым после малярии паразитарным заболеванием людей. Поражено около 120—150 млн. человек в 73 странах, расположенных в экваториальном, субэкваториальном, тропическом и субтропическом климатических поясах.
Читайте также:
