Читайте также:
- А- как экономические отношения;
- Административно-правовые нормы и правоотношения
- Алексеев. Правовой статус личности — это правовое положение человека, отражающее его фактическое состояние во взаимоотношениях с обществом и государством.
- Б) сохраняют силу, кроме случаев, когда в законе установлено, что его действие распространяется на отношения, возникшие из ранее заключенных договоров;
- Билет № 4 Конституционно-правовые отношения: понятие, особенности, виды
- Билет №17. Экономико-математические модели в компьютерном управлении.
- Билет №27 Сетевые модели. Детерминированные, вероятностные модели.
- Билет №32. Критический путь в сетевой модели. Минимизация общей продолжительности проекта с учетом трудовых ресурсов.
- Блоковая модель документа. Содержимое блока, рамки, поля и отступы. Блочные и строчные теги как элементы блоковой модели. Свойства display, overflow.
- Бюджетные правоотношения и нормы бюджетного права.
|
Хищник – жертва.
Некоторые животные способны менять окраску в зависимости от цвета фона. Например, головоногие моллюски, некоторые ракообразные, рыбы, амфибии, рептилии. Экспериментально показано, что из всех рыб, выживших в водоёме, где охотился пингвин, 66% составили особи, окрашенные соответственно фону.
Взаимодействия хищников и их жертв очень динамичны и взаимообусловлены. Это особенно ярко выражается в проявлениях адаптивного поведения. Наблюдения за головастиками американской жабы Bufo americans показали, что в присутствии хищных личинок стрекоз они снижают активность, изменяется и их распределение в пространстве. Например, у коловраток Brahionus calchforus в присутствии хищной коловратки Asplanchna развиваются длинные боковые выросты панциря, которые препятствуют заглатыванию их. Образование этих шпор индуцируется веществом выделяемым хищником и уменьшением пищи.
Паразит – хозяин.
Паразитизм – это форма взаимоотношений двух различных видов организмов, носящих онтогенетический характер, при которой организмы одного вида (паразита, потребителя) живут за счёт питательных веществ или тканей организма другого вида (хозяина) в течение определённого времени. Паразитизм известен на всех уровнях организации живого, начиная с вирусов и бактерий и заканчивая высокоорганизованными животными.
Распространение паразитизма. Чем сложнее устроен организм, тем выше эффективность его гомеостазирующих систем и поэтому больше благоприятных возможностей он предоставляет в качестве среды обитания. Чем организм совершеннее, тем меньше становится для него потребность использовать благоприятные условия в другом организме. Это объясняет, почему явление паразитизма наиболее представлено среди микроорганизмов и примитивных многоклеточных. Число видов, ведущих паразитический образ жизни, резко падает у высокоорганизованных организмов. У высших животных встречается наиболее обильное разнообразие населения паразитов.
Формы паразитизма и взаимные адаптации паразитов и их хозяев.
· Эктопаразитизм (клещи, напр. таёжный клещ, собачий клещ; блохи);
· Эндопаразитизм (паразитические черви, напр. печёночный сосальщик, широкий лентец).
По степени тесноты связей паразита и хозяина выделяют 2 формы:
1. Облигатный – ведёт только паразитический образ жизни, не выживает без обязательной связи с хозяином (все черви паразиты, напр. Taeniarhynchus saginatus – бычий цепень). Характеризуется наиболее полным комплексом приспособлений к использованию организма хозяина как среды.
2. Факультативный – ведут свободный образ жизни и лишь при особых условиях переходят к паразитическому существованию. Например, Armillaria mellea – опёнок. Их связь исключительно трофическая. Морфофизиологические и биологические адаптации к паразитизму выражены слабо.
Адаптации парзитов: среда в виде живого и подвижного организма требует формирования способов фиксации в ней: зацепки, крючья, присоски, видоизменения формы тела. Эктопаразиты имеют сплющенное тело. У эндопаразитов утрачиваются органы, которые не нужны им в среде хозяина.
Математические модели.
В естественных условиях (в гетерогенной среде) возникает следующая временная и причинно-следственная цепь: размножение жертвы→размножение хищника→резкое сокращение численности жертвы→падение численности хищника→размножение жертвы и т.д.
Волны жизни хищника и жертвы следуют друг за другом с постоянным сдвигом по фазе, и в среднем численность как хищника так и жертвы остаётся примерно на одном уровне. Длительность периода зависит от скорости роста обоих видов, а так же от исходных параметров. Для популяции жертвы влияние хищника положительно, т.к. её чрезмерное размножение привело бы к краху её численности. В свою очередь, все механизмы, препятствующие полному истреблению жертвы, способствуют сохранению пищевой базы хищника.
![]()
ГРАФИКИ. Взаимоотношения между хищником и жертвой: А – в гомогенной среде инфузория Dinidium nasatum погибает от голода после поедания жертвы; Б – в гетерогенной среде – жертва частично может укрыться от хищника, и он вымирает; В – естественные колебания численности хищника и жертвы ( по Гаузе).
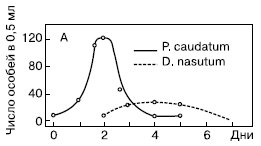
Рис. 3. Взаимодействие между инфузорией дидиниум и инфузорией туфелькой в однородной среде (количество особей в 0,5 см3)
![]()
Рис. 5. Взаимодействие между инфузорией дидиниум и инфузорией туфелькой в неоднородной среде (количество особей в 0,5 см)
![]()
Рис. 6. Взаимодействие между инфузорией дидиниум и инфузорией туфелькой в неоднородной среде при наличии иммиграций (1 инфузория туфелька + 1 дидиниум)
Эксперименты Гаузе.
Жертва – инфузория туфелька Paramecium caudatum.
Хищник – инфузория дидиниум Didinium nasutum.
В качестве пищи для инфузории туфельки Гаузе использовал культуру сенной палочки (Bacillus subtilis).
Первая серия опытов была поставлена Гаузе в так называемой однородной среде. Для этого использовалась разведенная бактериальная культура без осадка. Особенность биологии дидиниум заключается в том, что он не является активным охотником, а просто хватает все, что попадается ему на пути. Инфузория туфелька, ускользая от хищника, использует значительно бóльшую скорость передвижения, а также особые защитные образования – трихоцисты. При этом она никогда не прячется в убежища. Совокупность этих особенностей определила чистоту данной серии опытов и позволило Гаузе выделил процесс взаимодействия хищника и жертвы в чистом виде. Бактериальная культура без осадка разливалась в пробирки объемом 0,5 мл. В нее помещалось 5 особей инфузории туфельки, а через 2 суток вносили 3 особи хищной инфузории дидиниум. Практически сразу же после внесения хищников в культуру, численность жертвы начинала убывать, поскольку дидиниум поедает новую инфузорию туфельку через каждые 3 часа. Дидиниум интенсивно размножался, достигая максимума численности к 3-м – 4-м суткам опыта. На 5-е сутки дидиниум выел всех инфузорий туфелек, а на 6-е – все хищники погибли от голода.
При первой иммиграции в свободную среду дидиниум не нашел жертвы и погиб. После этого начался интенсивный рост инфузории туфельки. К моменту второй иммиграции концентрация инфузории туфельки в среде была уже очень велика, благодаря чему начался рост популяции дидиниум. Третья иммиграция пришлась на момент интенсивного уничтожения жертв хищником, и потому не внесла существенных изменений в общую картину. К четвертой иммиграции дидиниум выел всех инфузорий туфелек, после чего погиб сам. Внесенная в среду инфузория туфелька дала начало новому циклу роста жертв. В дальнейшем, такие изменения численности обоих видов регулярно повторялись.
Проанализировав результаты, полученные в ходе трех серий опытов, Гаузе пришел к заключению, что периодические колебания численности хищника и жертвы не являются свойством процесса взаимодействия между ними, как это предполагали математики. Вероятно, они возникают в результате постоянного вмешательства извне (убежища для жертв, сохранение хищников путем инцистирования, иммиграция и эмиграция, колебания факторов среды и т. д.). Именно поэтому в естественных условиях природа периодических колебаний численности находящихся в непосредственном взаимодействии видов может быть весьма сложной.
Дата добавления: 2015-04-18 ; просмотров: 29 ; Нарушение авторских прав
Аннотация научной статьи по сельскому хозяйству, лесному хозяйству, рыбному хозяйству, автор научной работы — Голубев А. В., Денисова Н. Б., Рубцов В. В.
Модели являются частью обширных программ биологической борьбы, разработанных для управления системой лес–насекомое. Существует много моделей паразит–хозяин , они построены на разных принципах и имеют различные параметры. Ф.Н. Семевский предложил унифицировать эти параметры на основе динамической системы. На примере непарного шелкопряда рассматривается его плотность и доля паразитированных тахинами гусениц. Установлено, что смертность, вызванная паразитами, при колебаниях численности хозяина имеет тенденцию сохранять свое значение. Таким образом, эмпирический факт относительного постоянства паразитизма подтверждает теоретические положения Ф.Н. Семевского.
Похожие темы научных работ по сельскому хозяйству, лесному хозяйству, рыбному хозяйству , автор научной работы — Голубев А. В., Денисова Н. Б., Рубцов В. В.
Models are part of an extensive program of biological control system designed to control forest insects. There are many models of host-parasite , they are based on different principles and have different parameters. F.N. Semevskii proposed combination of these parameters on the basis of a dynamic system. On the example of the gypsy moth is considered the density and the proportion of larvae that are infected tahini. It turns out that the mortality rate of parasites tends to constancy while the number of host varies. Thus, the empirical fact of the relative constancy of parasitism confirms theoretical position FN. Semevsky
МОДЕЛИ ПАРАЗИТ-ХОЗЯИН ДЛЯ ПРИНЯТИЯ РЕШЕНИЙ О ЦЕЛЕСООБРАЗНОСТИ БИОЛОГИЧЕСКОЙ БОРЬБЫ
A. В. ГОЛУБЕВ, проф. каф. экологии и защиты леса МГУЛ, д-р биол. наук,
Н.Б. ДЕНИСОВА, доц. каф. экологии и защиты леса МГУЛ канд. биол. наук,
B. В. РУБЦОВ, проф. Института лесоведения РАН, д-р биол. наук
Модели паразит-хозяин являются частью обширных программ биологической борьбы, разработанных для управления системой лес-насекомое [4].
В настоящее время имеется большое число моделей, построенных на различных принципах [8]. Это случайных поиск и постоянная поисковая эффективность; случайный поиск, поисковая эффективность зависит от плотности паразита; случайный поиск, поисковая эффективность зависит от плотности хозяина и паразита; неслучайный поиск и постоянная поисковая эффективность; неслучайный поиск, поисковая эффективность зависит от плотности паразита.
Проанализируем классическую модель Никольсона-Бейли. В оригинале модель выглядит следующим образом [8].
P-i = Nt(1 - exP(-aPX (1)
где Nt и - плотность популяции хозяина в год
Pt и Pt+1 - плотность популяции паразита в год t и t+1;
а - площадь, которую обследует самка за всю свою жизнь;
F - норма увеличения хозяина.
Заметим, что F не обязательно средняя плодовитость на самку, предположим, вид хозяина имеет среднюю плодовитость 100 яиц на самку. В этом случае F = 100, если только нет смертностей от различных факторов. Если же суммарная величина независимых смертностей составляет 90 %, F = 10, если смертность равна 95 %, F = 5 и т.д. [8].
Поисковую площадь можно вычислить из соотношения [6]
где а - обследуемая площадь;
y - плотность популяции самок паразито-идов;
P(y) - вероятность того, что определенная самка хозяина избежит заражения. Модель (1) порождает колебания с возрастающей амплитудой, однако ее практическое применение дает точные результаты [8].
Поскольку моделей паразит-хозяин оказалось много и все они построены на разных принципах и имеют различные параметры, Ф.Н. Семевский (личное сообщение) предложил унифицировать эти параметры на основе динамической системы
где х и у - соответственно численность паразита и хозяина;
а - среднее число паразитов, вышедших из одного зараженного хозяина;
к - коэффициент естественного воспроизводства хозяина.
Частными случаями этой модели являются модели Никольсона-Бейли и Хассела-Варли [1].
Однако подбор реальных параметров моделей и их экстраполяция встретили определенные трудности и от такого подхода было решено отказаться.
Ф.Н. Семевский [2] в соответствии с развитием им теоретическим представлениям поведения хозяина и паразитоида приписывается оптимальность (максимальное значение математического ожидания логарифма коэффициента размножения особи), целесообразность. В результате максимизируется средняя геометрическая коэффициента размножения особей. При этом модель Никольсона-Бейли дополняется системой уравнений, определяющих поверхность Ме-тью-Кермаки (любая адаптация развивается за счет совокупности прочих адаптаций) и задающих связь между адаптивными параметрами хозяина и паразитоида. После всех
ЛЕСНОЙ ВЕСТНИК 3/2013
Плотность популяции непарного шелкопряда и зараженных гусениц тахиной Parasetigena sylvestris
Участок Количество гусениц 1-го возраста на 100 сырой листвы / % паразитизма по годам
1991 1992 1993 1994
1 4,21/53,4 1,0/26,4 7,56/15,4 33,1/18,6
2 2,52/53,7 0,31/26,0 7,07/14,0 27,54/19,0
3 5,86/54,3 0,64/26,4 5,31/14,9 17,4/19,3
4 9,39/53,0 0,39/27,4 6,47/14,1 28,8/19,1
Контроль 1 25,4/23,7 3,01/24,7 6,97/14,8 30,1/18,9
Контроль 2 35,5/23,4 2,46/26,6 4,71/13,9 20,9/19,4
Прогноз и реализация движения численности непарного шелкопряда и зараженность гусениц тахиной
Прогноз на 1995 г. Реализация на 1995 г. Ошибка п рогноза, %
Хозяин Паразит Хозяин Паразит Хозяин Паразит
6,94 15,27 33,12 18,6 79 17,7
6,58 15,04 27,54 19,0 76 20,8
5,12 14,45 17,46 19,3 71 25,1
6,07 14,82 28,8 19,1 79 22,4
преобразований модель выглядит следующим образом [3]
где у и х - соответственно плотность популяции хозяина и паразита; b - коэффициент в рамках модели определяет постоянную смертность, вызванную паразитоидами;
У - средняя многолетняя плотность популяции хозяина.
Полученная модель оказалась глобально устойчивой [3], но ее предсказующая точность относительно удовлетворительная.
Рассмотрим данные по непарному шелкопряду и зараженность его гусениц тахиной Parasetigena sylvestris, полученные в дубовых насаждениях Самарской области [5] (табл. 1).
Из данных таблицы видно, что смертность, вызванная тахиной, при колебаниях численности хозяина имеет тенденцию сохранять свое значение. Исключением является только 1991 г, когда в результате обработки участков инсектицидами численность хозяина резко упала, и паразитизм по сравнению с контролем увеличился более чем в 2 раза. На следующий год общая тенденция восстановилась.
Таким образом, эмпирический факт относительного постоянства паразитизма подтверждает теоретические положения, высказанные [3].
Прогноз по модели 4 и реализация движения численности непарного шелкопряда и зараженность гусениц тахиной представлены в табл. 2.
Как и следовало ожидать, паразитизм мало варьирует по годам и модель дает достаточно удовлетворительное приближение к действительности. Наоборот, при прогнозировании плотности непарного шелкопряда модель обладает неудовлетворительной точностью.
Согласно [4] смертность, вызванная специализированными паразитами, при колебаниях численности имеет тенденцию сохранять свое значение; общая смертность, вызванная паразитами (специализированными и неспециализированными), еще более постоянна. Например [9] приводит такую модель для общего паразитизма гусениц старших возрастов листовертки-почкоеда
где p - доля паразитировенных особей;
N - плотность гусениц.
Таким образом, при анализе реальных процессов взаимодействия паразит-хозяин
ЛЕСНОЙ ВЕСТНИК 3/2013
можно предложить достаточно общий практический метод определения доли паразити-рованных особей. Если в этом году паразитизм был равен 30 %, то на следующий год он будет такой же.
1. Антоновкий, М.Я. Математический анализ некоторых эколого-экономических моделей / М.Я. Антоновкий, М.М. Семенов // Всесторонний анализ окружающей природной среды. - Л.: Гидрометео-издат, 1976. - С. 294-307.
2. Корзухин, М.Д. Синэкология леса. / М.Д. Корзу-хин, Ф.Н. Семевский. - СбП.: Гидрометеоиздат, 1992. - 192 с.
3. Семевский, Ф.Н. Динамика численности животных, растений и человека / Ф.Н. Семевский. - Екатеринбург, 2007. - 200 с.
4. Семевский, Ф.Н. Математическое моделирование экологических процессов / Ф.Н. Семевский, С.М. Семенов. - Л.: Гидрометеоиздат, 1982. - 280 с
5. Сергеева, Ю.А. Адаптации и устойчивость системы паразит-хозяин./ Ю.А. Сергеева, А.В. Голубев // Экология, мониторинг и рациональное природопользование: Научные труды МГУЛ. - М.: МГУЛ, 1996. - С. 64-70.
6. Смит, Дж.М. Модели в экологии / Дж.М. Смит. - М.: Мир, 1976. - 184 с.
7. Холинг, К.С. (ред.) Экологические системы. Адаптивная оценка и управление / К.С. Холинг. - М.: Мир, 1981. - 397 с.
8. Hassel M.P., May R.M. Stability in insect host-parasite Models. // J/ Anim. Ecol., 1973, v. 42, 3. - P. 693726/
9. Jones D.D. The budworm site Model/ //Proceed. Confer. In Pest Management IIASA H-79-42 Gb, 1979, v. 4. - 155p/
очагов типографа в брянской области в период кульминации размножения
В.С. КЛЮЕВ, асп. каф. лесозащиты и охотоведенияБрГИТА,
В.П. ШЕЛУХО, проф. каф. лесозащиты и охотоведения БрГИТА, д-р с.-х. наук
Резкому росту численности и активности типографа способствовала экстремальная жара и засуха лета 2010 г., которая, с одной стороны, способствовала увеличению числа поколений короеда, развивающихся в течение года, а с другой стороны - привела к значительному ослаблению ельников, особенно на хорошо дренированных почвах и возвышенных элементах рельефа.
Типограф относительно свето- и теплолюбив, предпочитает среднеполнотные
Наши исследования проводились в еловых насаждениях на территории Брянской области, которые занимают площадь 111,7 тыс. га. Средний возраст ельников составляет 42 года, средняя полнота - 0,74, средний класс бонитета находится на уровне 1,2.
Информация
Добавить в ЗАКЛАДКИ
| Поделиться: |
Паразит хозяин, система
Равновесие в системах хищник—жертва и паразит—хозяин не возникает автоматически, а устанавливается на протяжении длительного времени (много тысяч и даже миллионов лет). За такое время виды, взаимодействующие подобным образом, адаптируются друг к другу и к среде обитания так, что естественные враги не уничтожают полностью популяцию жертвы.[ . ]
Биологическое равновесие д системе паразит — хозяин — необходимое условие длительного, теоретически вечного существования биоценозов и сохранения их видового разнообразия. Если бы паразиты уничтожили своих хозяев, то они лишились бы своего места обитания и источника пищи. Гибель хозяев неизбежно привела бы к вымиранию паразитов. Однако паразиты могут влиять на хозяев и позитивно. Так, паразитирование пиявок на рыбах приводит к повышению эффективности использования хозяином корма. При взаимодействии паразит — хозяин паразитизм благоприятно влияет на популяции и паразита, и хозяина.[ . ]
Биологическое равновесие д системе паразит — хозяин — необходимое условие длительного, теоретически вечного существования биоценозов и сохранения их видового разнообразия. Если бы паразиты уничтожили своих хозяев, то они лишились бы своего места обитания и источника пищи. Гибель хозяев неизбежно привела бы к вымиранию паразитов. Однако паразиты могут влиять на хозяев и позитивно. Так, паразитирование пиявок на рыбах приводит к повышению эффективности использования хозяином корма. При взаимодействии паразит — хозяин паразитизм благоприятно влияет на популяции и паразита, и хозяина.[ . ]
Биологическое равновесие д системе паразит — хозяин — необходимое условие длительного, теоретически вечного существования биоценозов и сохранения их видового разнообразия. Если бы паразиты уничтожили своих хозяев, то они лишились бы своего места обитания и источника пищи. Гибель хозяев неизбежно привела бы к вымиранию паразитов. Однако паразиты могут влиять на хозяев и позитивно. Так, паразитирование пиявок на рыбах приводит к повышению эффективности использования хозяином корма. При взаимодействии паразит — хозяин паразитизм благоприятно влияет на популяции и паразита, и хозяина.[ . ]
Другая причина негативных изменений в системе паразит — хозяин — сортовая унификация семян культурных растений, предназначенных для посева. При унификации агротехники, методов посева культур, уборки, хранения и переработки урожая предпочтение отдается определенному сорту растений. Но устойчивость любого сорта к возбудителю болезни относительна. Возбудитель болезни рано или поздно преодолевает устойчивость растения к его патогенному воздействию. Устойчивость культурных растений того или иного сорта к грибному заболеванию — стеблевой ржавчине — обычно не превышает пяти лет.[ . ]
Большинство исследователей считает, что у ремнецветниковых корневой паразитизм первичен, а стеблевой — вторичен. Одна группа стеблевых паразитов образует корнеподобные органы (эпикортикалыше корни), растущие по поверхности ветвей растения-хозяина. Эти сильно преобразованные органы морфологически, вероятно, гомологичны придаточным корням. Через определенные интервалы на них образуются погружающиеся в ткань растения-хозяина гаустории, с помощью которых паразит закрепляется на ветви и получает из нее воду и питательные вещества. Такой способ паразитирования характерен, например, для вида струтантуса (Slrutlianlhus) и представителей некоторых других родов. У более специализированных паразитов мощная гаусториаль-ная система в виде длинных ветвящихся тяжей или аморфных образований развивается внутри стеблей растения-хозяина. На их поверхности появляются только побеги и соцветия паразита, возникающие в разных точках из внутренних тканей. Такой глубокий паразитизм характерен для некоторых видов ремнецветника и пситтакантуса (Psittacantlms). Почти все ремнецветниковые размножаются вегетативно. Гаустории у этих паразитов образуются из всех вегетативных органов — корней, стеблей и даже листьев. У тропического американского вида струтантуса округлого (S. orbicularis) свисающие тонкие молодые ветви имеют крепкие загнутые листья с крючковатой вершиной. Когда такой лист зацепляется за ветвь дерева, черешок закручивается вокруг нее, и вскоре в этом узле образуются эпикортикалыше органы, охватывающие ветвь хозяина и внедряющие в нее свои, гаустории. В условиях тропического леса с его обильной растительностью часто наблюдается прикрепление одного паразита сразу к нескольким растениям-хо-зяевам и даже к другим паразитным растениям.[ . ]
Поражение корней питиевыми грибами носит название корнееда или корневой гнили. Корневой гнилью поражаются всходы сахарной свеклы, хлопчатника, люцерны и деревьев (рис. 29,2). Особенно сильно корнеед развивается в годы с холодной и влажной весной, когда корневая система развивается медленно, а отдельные участки корней вследствие недостатка воздуха в переувлажненной почве отмирают. Мертвые клетки не способны к активной физиологической защите и легко заселяются грибами, служа воротами инфекции. Поселившись на отмерших участках корней, паразит питается там и выделяет токсические вещества, которые убивают прилегающие к мертвым живые участки. Таким образом осуществляется продвижение паразита по ткани хозяина. Подобные виды питиевых грибов, так же как и паразиты водорослей, широко специализированы и могут заражать десятки и сотни видов высших растений из разных родов и семейств.[ . ]
Еще больший вред приносят ленточные черви — свиной и бычий цепень (солитер), большой лентец, эхинококк. Циклы их развития достаточно сложны. Яйца этих червей может съесть домашнее животное с травой или с отбросами, как это делают свиньи. Яйца могут быть смыты в водоем и попасть в кишечник рыбы. В кишечнике промежуточного хозяина из яйца развивается личинка с тремя парами крючков. Пробуравливая стенки кишечника, она попадает в кровь и с током крови заносится в мышцы. В мышцах личинка превращается в покоящуюся стадию — финну и может просуществовать в таком виде сколь угодно долго. Человек заражается, если съедает плохо проваренное или прожаренное финнозное мясо. Пищеварительная система цепня редуцирована, питательные вещества он всасывает всей поверхностью тела. Паразит присасывается к стенкам кишечника. Червь быстро растет и делится, образуя членики, его длина может достигать 2—3 м. Задние членики созревают раньше и заполняются яйцами. Членики отрываются, вынося наружу тысячи яиц. Солитер особенно опасен для человека, когда в его организм попадают яйца цепня. Тогда в мышцах, а иногда и в мозге образуются финны.[ . ]
Сопряженная эволюция, или коэволюция, рассматриваемая нами на внутри- и межвидовом уровнях, отличается тем, что при ней обмен генетической информацией минимален. На уровне сообществ можно рассматривать селективные воздействия между группами организмов, находящихся в экологическом взаимодействии: растения и растительноядные животные, крупные организмы и мелкие симбионты, паразит — хозяин, хищник — жертва и т.д. На уровне биотических сообществ коэволюция — это сближение в результате взаимной адаптации двух взаимосвязанных эволюционирующих систем, когда изменение, произошедшее в одной системе, инициирует такое изменение в другой, которое не приводит к нежелательным или тем более неприемлемым для первой системы последствиям (Данилов-Данильян, Лосев, 2000).[ . ]
Еще одна причина возникновения эпифитотий — повышение пищевой ценности культурных растений как звена трофической цепи. С помощью агротехнических приемов, селекции и семеноводства, других методов проводят мероприятия по улучшению пищевых качеств растений. Например, выведены сорта злаковых растений с высоким содержанием аминокислот, в том числе незаменимых (в частности, лизина). Но все, что полезно для человека и животных, не менее полезно и для возбудителей болезней, и для вредителей растений. Растения с высокими пищевыми качествами обеспечивают паразитов-патогенов-вредителей полноценной высококалорийной пищей (кормом). В агроценозе вредители и возбудители болезней растений находятся в условиях изобилия высококачественной пищи. Энергетические затраты паразитов на добывание пищи, удовлетворение потребностей организма в питательных веществах, минеральных солях и витаминах значительно ниже, чем в дикой природе. Условия для роста, развития и размножения паразитов улучшаются, биологическое равновесие в системе паразит — хозяин смещается в пользу патогена.[ . ]
Надорганизменные системы, которые изучает экология — популяции, биоценозы, экосистемы — чрезвычайно сложны. В них возникает множество взаимосвязей, сила и постоянство которых непрерывно меняются. Одни и те же внешние воздействия могут привести к различным, иногда прямо противоположным результатам, в зависимости от того, в каком состоянии находилась система в момент воздействия.
Предвидеть ответные реакции системы на действие конкретных факторов можно лишь через сложный анализ существующих в ней количественных взаимоотношений и закономерностей. В экологии поэтому широкое распространение получил метод математического моделирования как средство изучения и прогнозирования природных процессов.
Суть метода заключается в том, что с помощью математических символов строится абстрактное упрощенное подобие изучаемой системы. Затем, меняя значение отдельных параметров, исследуют, как поведет себя данная искусственная система, т. е. как изменится конечный результат.
В построении математических моделей сложных процессов выделяются следующие этапы.
1. Прежде всего, те реальные явления, которые хотят смоделировать, должны быть тщательно изучены: выявлены главные компоненты и установлены законы, определяющие характер взаимодействия между ними. Если неясно, как связаны между собой реальные объекты, построение адекватной модели невозможно. На этом этапе должны быть сформулированы те вопросы, ответ на которые должна дать модель. Прежде чем строить математическую модель природного явления, надо иметь гипотезу о его течении.
2. Разрабатывается математическая теория, описывающая изучаемые процессы с необходимой детальностью. На ее основе строится модель в виде системы абстрактных взаимодействий. Установленные законы должны быть облечены в точную математическую форму. Конкретные модели могут быть представлены в аналитической форме (системой аналитических уравнений) или в виде логической схемы машинной программы. Модель природного
явления есть строгое математическое выражение сформулированной гипотезы.
3. Проверка модели —расчет на основе модели и сличение результатов с действительностью. При этом проверяется правильность сформулированной гипотезы. При значительном расхождении сведений модель отвергают или совершенствуют. При согласованности результатов модели используют для прогноза, вводя в них различные исходные параметры.
Следует, однако, отметить, что сама по себе математическая модель не может служить абсолютным доказательством правильности той или иной гипотезы, так как может оказаться, что разные гипотезы приводят к сходным результатам, но она служит . одним из путей анализа реальности.
Моделируют различные по характеру процессы, происходящие в реальной среде, как, например, отдельные типы экологических взаимодействий хищник — жертва, паразит — хозяин, конкурентные отношения, мутуализм и др. Математическими моделями описываются и проверяются разные варианты динамики численности популяций, продукционные процессы в экосистемах, условия стабилизации сообществ, ход восстановления систем при разных формах нарушений и многие другие явления. Сами методы математического моделирования биологических систем развиваются, совершенствуются и разнообразятся.
Например, одну из простейших математических моделей для системы паразит — хозяин в динамике численности насекомых разработал в 1925″г. А. Лотки, который вывел следующие уравнения:
![]()
где Ni — численность популяции хозяина; N2 — численность популяции паразита; г — удельная скорость увеличения популяции хозяина; d2 —удельная скорость гибели популяции паразита; рхл р2 — константы. График процесса паразитической инвазии, построенный по таким уравнениям, обнаруживает, что в результате взаимодействия двух видов должны возникать осцилляции (колг-бания) с постоянной амплитудой, которая зависит от соотношения между скоростями увеличения численности двух видов.
В период, когда были сделаны эти расчеты, экологи вели поиск причин циклических колебаний численности, которые были обнаружены к тому времени у ряда видов. Делались попытки отыскать внешние факторы (космические, солнечные, атмосферные), ответственные за периодические изменения популяций. Модели А. Лотки и В. Вольтерра позволили выдвинуть идею, что периодический колебательный режим в популяциях может возникнуть в результате межвидовых отношений и без внешнего периодического воздействия. Эта идея оказалась плодотворной для дальнейшего развития теории динамики численности популяций. Однако сама модель являлась не адекватной, т. е. не описывала действительность, так как в природе практически не обнаруживаются подобные непрерывные осцилляции с постоянной амплитудой у пар видов, связанных по типу хищник — жертва или паразит — хозяин.
В 1933 г. А. Никольсон, несколько усложнив математическую модель Лотки и введя в систему дополнительных хозяев и паразитов, показал, что это ослабляет осцилляции. В 1936 г. A. >i. Кол мо гор о в разработал новые подходы и описал также возможности устойчивого стационарного состояния системы взаимодействующих через трофические связи видов. Позднее для систем хищник — жертва, паразит — хозяин было предложено множество других моделей. С введением в модели дополнительных параметров сильно усложняется математический аппарат и техника расчетов. Многие из этих ограничений позволило снять использование электронно-вычислительных машин.
![]()
![]()
С 60-х годов появляется множество работ, посвященных математическому анализу с помощью ЭВМ взаимоотношений хищник — жертва и паразит — хозяин. Экспериментирование с моделями на вычислительных машинах открыло широкие возможности для поиска стратегий управления биологическими системами. Моделирование на ЭВМ позволяет также совершенствовать систему сбора исходных сведений. Так, если модель содержит нереалистические предположения, то картина на выходе ЭВМ позволяет понять, как надо упорядочить эксперименты и наблюдения для получения необходимой количественной информации.
Модели хищник — жертва играют большую роль в планировании рыбного, китобойного, охотничьего промыслов, так как изъятие человеком части популяции диких животных с экологических позиций является аналогом природного хищничества. Предельная степень эксплуатации, которую может выдержать популяция, различна у разных видов. Важно вовремя заметить симптомы, свидетельствующие, что изъятие из популяции приближается к предельно допустимому уровню, после которого может быть нарушена ее воспроизводительная способность.
Например, по результатам машинных экспериментов со статистикой китобойного промысла в 60-х годах выявлены показатели допустимых масштабов добычи и симптомы гибельной эксплуатации популяции синих китов. Если популяция эксплуатируется интенсивно, но не чрезмерно, то в моделях обнаруживается уменьшение размеров и среднего возраста особей, кривые выживания изменяются, но не настолько, чтобы нарушалась плодовитость стада в целом. В реальной действительности были обнаружены предсказанные моделями симптомы гибельной эксплуатации китового стада — сокращение доли беременных самок, сильные изменения кривых выживания, уменьшение размеров уловов на единицу промыслового усилия, неспособность популяции быстро восстановить численность после прекращения промысла. Синих китов осталось так мало, что несмотря на международный запрет их добычи, принятый в 1967 г., поголовье остается на низком уровне и животные внесены в Красную книгу.
Моделирование трофических связей имеет большое значение для решения проблем борьбы с вредителями, регуляции численности популяций, стабилизации сообществ.
Математическое моделирование широко применяется при решении экологических проблем, связанных с антропогенными воздействиями на природную среду. В современных математических моделях выделяют тактические и стратегические модели. Тактические модели экосистем и популяций служат для экологического прогнозирования их состояния, в том числе при разного рода экзогенных воздействиях. Стратегические модели строят в основном с исследовательскими целями, для вскрытия общих законов функционирования биологических систем, таких, как стабильность, разнообразие, устойчивость к воздействиям, способность возвращаться в исходное состояние. В задачи стратегических моделей входит изучить с помощью ЭВМ последствия разных стратегий управления экосистемами, чтобы иметь возможность выбрать оптимальную.
Модели, которые описывают взаимодействие общества и природы и в которых учитывают не только экологические, но и экономические, демографические и социальные показатели, называют эколого-экономическими моделями. Такие модели разрабатывают для долгосрочного прогнозирования экономического роста и общей оценки влияния человеческой деятельности на природную среду.
Читайте также:
 Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу. Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу.
Copyright © Иммунитет и инфекции
|