
Лактозный оперон у кишечной палочки

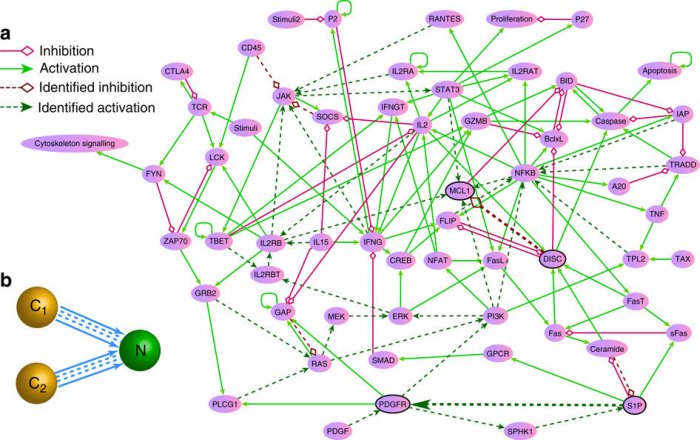
Упрощенная схема генной сети, регулирующей деление клеток человека
Как устроена внутриклеточная автоматика?
Где есть сигналы, там будут наводки — сигналы не на своем месте и шумы — случайная фигня, прилипающая к сигналам. В электронике приход сигналов по адресу обеспечивается проводами и изоляцией. Наводки возникают из-за паразитных емкостных и индуктивных связей между проводниками. В клетке бактерии ее содержимое более-менее равномерно перемешано, и любой химический сигнал (а их тысячи) доступен в любой ее точке. Клетки животных и растений разделены на отсеки с разным химическим составом, но типов этих отсеков меньше десятка и в каждом есть сотни и тысячи разных веществ.

Что конкретно регулируется в живой клетке?
Клетку можно рассматривать как завод, который производит все необходимое ему оборудование и может собирать из него второй такой же завод. Основным оборудованием являются рибосомы — станки с ЧПУ, собирающие все белки. Структура собираемого белка записана на ленте — матричной РНК (мРНК). Рибосома движется по мРНК и по ее инструкции собирает цепочку нового белка из отдельных звеньев — аминокислот. Выходящая из рибосомы белковая цепочка сворачивается в компактный клубок зрелого белка, который начинает работать. Белки могут ускорять химические реакции (ферменты), а так же заниматься транспортом веществ, передачей сигналов, защитой и многими другими делами.
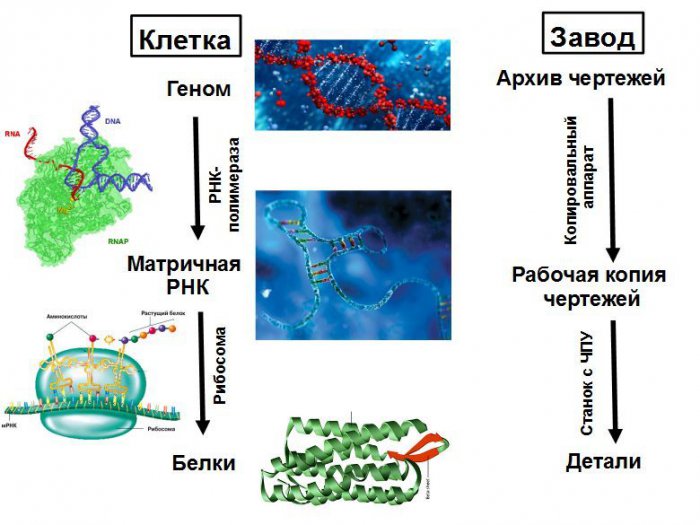
мРНК образуются как копии отдельных участков клеточной ДНК. ДНК можно сравнить с архивом технологической документации в кабинете главного инженера, а мРНК — с копиями чертежей, которые выдаются в цех. Копировальный аппарат, создающий эти копии — это тоже белок, который называется РНК-полимераза. Она может садиться на ДНК и начинать ее копирование не везде, а только на специальных участках ДНК — промоторах, которые находятся перед каждым геном или группой работающих вместе генов.
Включение и выключение генов определяется в основном на этапе посадки РНК-полимеразы на промотор. Если РНК-полимераза легко садится на промотор гена, то с него делается много мРНК-копий и рибосомы производят много молекул белка, кодируемого этим геном — он будет включен. Если что-то помешает посадке РНК-полимеразы, то ген окажется выключен. То есть, включение и выключение генов происходит на этапе взаимодействия РНК-полимеразы и промотора. Различные другие молекулы могут помогать или мешать их связыванию. Эти другие молекулы, в первую очередь специальные регуляторные белки, вместе с промотором и соседними участками ДНК образуют логический элемент, который может интегрировать несколько разных входных сигналов.
Логические элементы на ДНК и белках
Пожалуй, самый изученный пример генного переключателя — лактозный оперон кишечной палочки. Французы Жакоб и Моно за его исследования были удостоены Нобелевской премии в 1965 году. Кишечная палочка, как понятно из ее названия, живет в кишечнике. Это может быть кишечник самых разных животных, от пчелы до человека. Ей там приходится питаться тем, что попалось хозяину, поэтому она может есть много разных питательных веществ.
Кишечная палочка под микроскопом и на чашке Петри
Кишечная палочка может расти, например, на питательной среде, состоящей только из глюкозы и минеральных солей и производить все необходимые ей аминокислоты и витамины. Вместо глюкозы могут быть и другие сахара (фруктоза, солодовый сахар — мальтоза, молочный сахар — лактоза и десяток других), а так же полисахариды, например крахмал.
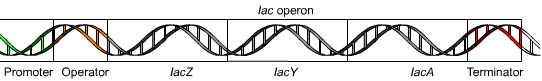
Схема лактозного оперона
Активность лактозного оперона управляется двумя сигналами. Первый сигнал, это, очевидно, концентрация лактозы. Если лактозы нет, то и ферменты ее усвоения не нужны. Второй сигнал чуть сложнее. В одном из экспериментов Жакоб и Моно растили кишечную палочку на среде, содержащей и лактозу, и с глюкозу. В таких условиях палочка сначала потребляет глюкозу, а лактозный оперон у нее выключен. Когда глюкоза кончается, рост бактерий приостанавливается минут на 15–20, а затем продолжается уже за счет лактозы. Во время паузы лактозный оперон включается. То есть, второй регуляторный вход выключает лактозный оперон, когда у клетки есть более доступные сахара, чем лактоза (усвоение глюкозы не требует никаких дополнительных ферментов).
Лактозный репрессор — это белок, который умеет связываться либо с лактозой, либо с определенной последовательностью ДНК. В геноме кишечной палочки есть две такие последовательности, обе в начале лактозного оперона, по обе стороны от промотора. Молекулы лактозного репрессора прочно слипаются друг с другом в группы по четыре белковых молекулы. Связывание такой четверки с ДНК закручивает нить ДНК в крутую петлю, в которой оказывается промотор. РНК-полимераза не может сесть на такой закрученный промотор и ген оказывается выключен. Если в клетке появляется лактоза, она связывается с лактозным репрессором и он отваливается от ДНК, освобождая промотор для РНК-полимеразы — оперон начинает работать. Так устроен регуляторный вход оперона по концентрации лактозы.
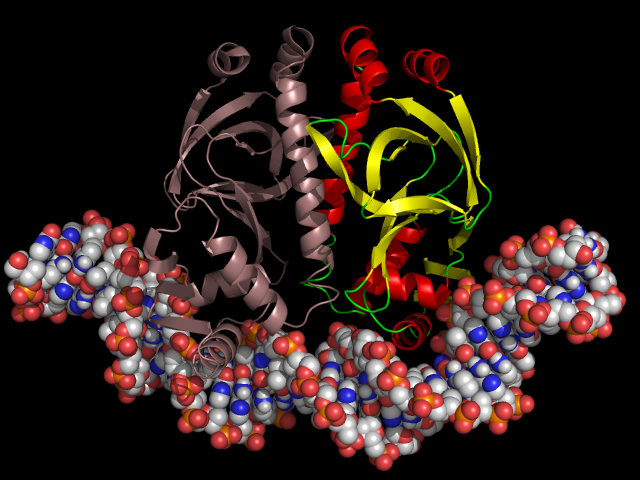
Катаболический активатор на ДНК
Катаболический активатор похожим способом реагирует на концентрацию цАМФ. Правда, есть несколько отличий в деталях:
— катаболический активатор садится на ДНК чуть сбоку от промотора и не перекрывает его. Наоборот, он усиливает связывание РНК-полимеразы с промотором и повышает активность оперона.
— катаболический активатор связывает цАМФ и ДНК. Без цАМФ он не может связаться с ДНК.
— катаболический активатор нужен для полной активности лактозного оперона, но и без него оперон все же будет работать примерно на 5% от полной активности. Лактозный репрессор же регулирует оперон в диапазоне от 0 до 100%.
— молекулы катаболического активатора соединяются в пары, а не в четверки. В следующем посте мы увидим, к чему приводит эта разница.
То есть, мы можем написать таблицу истинности для лактозного оперона:
Продолжение следует. В нем будут графики и матан.
| ← Предыдущая глава | Глава 3.10 | Следующая глава → |
| Регуляция работы гена: лактозный оперон | ||
Строение лактозного оперона [ править ]
Оперон — функциональная единица генома у прокариот, в состав которой входят гены, кодирующие совместно или последовательно работающие белки и часто объединенные под одним промотором.
Концепцию оперона для прокариот предложили в 1961 году французские ученые Франсуа Жакоб, Жан Моно и Андре Львов, за что получили Нобелевскую премию в 1965 году.
Характерным примером оперонной организации генома прокариот является лактозный оперон. Регуляция экспрессии генов метаболизма лактозы у кишечной палочки (Escherichia coli) была впервые описана в 1961 году учеными Ф. Жакобом и Ж. Моно.
Лактозный оперон (lac оперон) — полицистронный оперон бактерий, в состав которого входят гены, отвечающие за усвоение и расщепление лактозы.
Лактозный оперон состоит из промотора, оператора, трех структурных генов и терминатора. Иногда принимается, что в состав оперона входит также ген-регулятор, который кодирует белок-репрессор.
Промотор — последовательность нуклеотидов ДНК, узнаваемая РНК-полимеразой как стартовая площадка для начала специфической, или осмысленной, транскрипции. У прокариот все промоторы включают ряд похожих последовательностей нуклеотидов, важных для узнавания их РНК-полимеразой; в то же время разные промоторы отличаются друг от друга по другим последовательностям. Промотор асимметричен, что позволяет РНК-полимеразе начать транскрипцию в правильном направлении и указывает то, какая из двух цепей ДНК будет служить матрицей для синтеза РНК. Промотор может частично перекрываться или вовсе не перекрываться с оператором.
Оператор — это последовательность нуклеотидов ДНК, с которой связывается регуляторный белок — репрессор или активатор.
Структурные гены — это гены, кодирующие белки.
Структурные гены лактозного оперона — lacZ, lacY и lacA:
- lacZ кодирует фермент β-галактозидазу, которая расщепляет дисахарид лактозу на глюкозу и галактозу,
- lacY кодирует β-галактозид пермеазу, мембранный транспортный белок, который переносит лактозу внутрь клетки.
- lacA кодирует β-галактозид трансацетилазу, фермент, переносящий ацетилную группу от ацетил-КoA на бета-галактозиды.
Для усвоения лактозы необходимы только продукты генов lacZ и lacY, роль продукта гена lacA не ясна.

Регуляция работы лактозного оперона [ править ]
Бактериальная клетка синтезирует ферменты, принимающие участие в метаболизме лактозы, лишь в том случае, когда лактоза присутствует в окружающей среде и клетка испытывает недостаток глюкозы.
РНК-полимераза начинает транскрипцию с промотора, который в случае лактозного оперона перекрывается с оператором. В отсутствие или при низкой концентрации лактозы в клетке белок-репрессор, который является продуктом гена LacI, обратимо соединяется с оператором и препятствует транскрипции. Таким образом, в отсутствие лактозы в клетке ферменты для метаболизма лактозы не синтезируются.
Даже в случае, когда в плазматической мембране клетки отсутствует фермент β-галактозидпермеаза, лактоза из окружающей среды может попадать в клетку в небольших количествах. В клетке две молекулы лактозы связываются с белком-репрессором, что приводит к изменению его конформации и далее к отделению белка-репрессора от оператора. Теперь может осуществляться транскрипция генов лактозного оперона. При снижении концентрации лактозы новые порции белка-репрессора взаимодействуют с операторными последовательностями и препятствуют транскрипции. Данный механизм регуляции активности лактозного оперона называют позитивной индукцией. Веществом-индуктором служит лактоза; при её связывании с белков-репрессором происходит его отделение от оператора.
Итак, ферменты для усвоения лактозы синтезируются в клетке кишечной палочки при двух условиях: 1) наличие лактозы; 2) отсутствие глюкозы.
Вопрос 1 Мутации каких участков в ДНК кишечной палочки могут привести к появлению бактерий:
- Не способных усваивать лактозу?
- Синтезирующих ферменты для усвоения лактозы даже в её отсутствие?
- Способных усваивать лактозу при высокой концентрации глюкозы?
Биологический смысл [ править ]
Благодаря описанному механизму регуляции транскрипции генов, входящих в состав лактозного оперона, бактерии оптимизируют энергетические затраты, синтезируя ферменты метаболизма лактозы не постоянно, а лишь тогда, когда клетке это необходимо. Сходный механизм регуляции имеется у большинства прокариот; у эукариот он устроен значительно сложнее.
Регуляция работы лактозного оперона в зависимости от концентрации лактозы происходит по принципу отрицательной обратной связи: чем больше лактозы — тем больше ферментов для её усвоения (положительная прямая связь); чем больше ферментов — тем меньше лактозы, чем меньше лактозы — тем меньше производится ферментов (двойная отрицательная обратная связь).
Презентация была опубликована 6 лет назад пользователемБогдан Капитонов
Презентация на тему: " Лактозный оперон Подготовил: Проверил:. Введение Группы генов Строение гена Виды оперонов Лактозный оперон. Схема строения lac-оперона Структурные гены." — Транскрипт:
1 Лактозный оперон Подготовил: Проверил:
2 Введение Группы генов Строение гена Виды оперонов Лактозный оперон. Схема строения lac-оперона Структурные гены участвующие в метаболизме лактозы Негативная регуляция транскрипции lac-оперона Позитивная регуляция транскрипции lac-оперона Схема позитивной регуляции транскрипции лактозного оперона Транскрипция Вывод
3 Первым и главным элементом контроля реализации потока генетической информации на пути от ДНК к белку у всех организмов является контроль на уровне транскрипции. Регуляция транскрипции у прокариот обычно охватывает группу генов, кодирующих функционально родственные белки, участвующие в осуществлении связанных между собой химических превращений в клетке. Такими белками обычно являются ферменты. Группа согласовано регулируемых генов, кодирующих эти ферменты, называется опероном.
5 Группы генов по функциям Группы генов по функциям Структурные (конститутивные) (белки ферменты, гистоны, последовательность нуклеотидов в РНК) Гены –модуляторы: ингибиторы, интенсификаторы, интеграторы, модификаторы. Функциональные (индуцибельные) Гены – регуляторы, регулирующие работу структурных генов.
6 Структурная часть гена Промотор Начало транскрипции Старт-кодон Стоп - кодон Терминатор транскрипции Строение гена Строение гена Участок связывания фермента РНК- полимеразы (место начала транскрипции) Участок, кодирующий последовательность аминокислот в молекуле белка. Триплет ТАЦ в ДНК и АУГ в РНК иницирующий начало синтеза белка Триплеты – бессмысленные кодоны на которых завершается трансляция Участок, сигнализирующий о прекращении транскрипции
7 Оперон Оперон это тесно связанная последовательность структурных генов, определяющих синтез группы белков, которые участвуют в одной цепи биохимических преобразований. К наиболее хорошо изученным оперонам бактерий относят: Лактозныйlacоперон Лактозный (lac)-оперон Галактозныйgalоперон Галактозный (gal)-оперон Триптофановыйtrpоперон Триптофановый (trp)-оперон Рассмотрим механизмы регуляции активности генов на примере лактозного оперона кишечной палочки. Оперон Оперон это тесно связанная последовательность структурных генов, определяющих синтез группы белков, которые участвуют в одной цепи биохимических преобразований. К наиболее хорошо изученным оперонам бактерий относят: Лактозныйlacоперон Лактозный (lac)-оперон Галактозныйgalоперон Галактозный (gal)-оперон Триптофановыйtrpоперон Триптофановый (trp)-оперон Рассмотрим механизмы регуляции активности генов на примере лактозного оперона кишечной палочки.
8 Оперон Оперон – участок бактериальной хромосомы, включающий следующие участки ДНК: Р – промотор, О – оператор, Z, Y, А – структурные гены, Т –терминатор. (В состав других оперонов может входить до 10 структурных генов.) Промотор Промотор – место присоединения РНК- полимеразы. Оператор Оператор – участок оперона, к которому присоединяются белки- репрессоры или активаторы транскрипции. Структурныегены Структурные гены – гены кодирующие ферменты, участвующие в метаболизме лактозы. Терминатор Терминатор служит для отсоединения РНК-полимеразы после окончания синтеза иРНК, соответствующей ферментам Z, Y, А, необходимым для усвоения лактозы. Оперон Оперон – участок бактериальной хромосомы, включающий следующие участки ДНК: Р – промотор, О – оператор, Z, Y, А – структурные гены, Т –терминатор. (В состав других оперонов может входить до 10 структурных генов.) Промотор Промотор – место присоединения РНК- полимеразы. Оператор Оператор – участок оперона, к которому присоединяются белки- репрессоры или активаторы транскрипции. Структурныегены Структурные гены – гены кодирующие ферменты, участвующие в метаболизме лактозы. Терминатор Терминатор служит для отсоединения РНК-полимеразы после окончания синтеза иРНК, соответствующей ферментам Z, Y, А, необходимым для усвоения лактозы.
9 Регуляторная область Полицистронная единица транскрипции lac ZlacYlac A АктиваторПромотор Оператор Спейсеры Нетраслируемая 5 ʹ - последовательность Точка терминации транскрипции 3ʹ3ʹ 5ʹ5ʹ 5ʹ5ʹ 3ʹ3ʹ
10 Структурные гены участвующие в метаболизме лактозы Ген lac ZГен lac A Ген lac Y Кодирует фермент β- галактозидазу, расщепляющий β- галактозиды, в частности лактозу, с образованием моносахарида – глюкозы. Кодирует β- галактозидтрансферазу (пермеазу), фермент необходимый для проникновения β- галактозидов через клеточную мембрану. Кодирует фермент β- галактозидтрансацетил азу, функциональная роль которого до конца не выяснена.
11 Оператор – это область с которой взаимодействует белок-репрессор. Сам lac-репрессор является продуктом экспрессии соответствующего гена (i-гена) и содержится у E. coli в количестве не более 10 молекул на 1 клетку. Активная форма lac-репрессора представляет собой гомотетрамер, который связывается с зоной оператора и блокирует действие РНК-полимеразы. Такая негативная регуляция транскрипции наблюдается при достаточной концентрации глюкозы в клетке и в отсутствии избытка лактозы. Оператор – это область с которой взаимодействует белок-репрессор. Сам lac-репрессор является продуктом экспрессии соответствующего гена (i-гена) и содержится у E. coli в количестве не более 10 молекул на 1 клетку. Активная форма lac-репрессора представляет собой гомотетрамер, который связывается с зоной оператора и блокирует действие РНК-полимеразы. Такая негативная регуляция транскрипции наблюдается при достаточной концентрации глюкозы в клетке и в отсутствии избытка лактозы.
13 Ген-регулятор иРНК-полимераза Оператор Структурный ген транскрипция иРНК трансляция Лактоза действует как индуктор Комплекс индуктора с репрессором, неспособный связаться с оператором Глюкоза + галактоза β - галактозидаза фермент Схема позитивнойрегуляции транскрипции лактозногооперона Схема позитивной регуляции транскрипции лактозного оперона Лактоза репрессор Рассмотрим механизм регуляции синтеза белка на примере работы лактозного оперона молочнокислых бактерий
15 Работа оперона молочно-кислых бактерий происходит в результате деятельности белка репрессора и факторов внешней среды (наличие или отсутствие индуктора) Как происходит работа оперона молчно-кислых бактерий? Как действуют гены в работе оперона? Процесс расщепления лактозы происходит в ходе взаимодействия генов, входящих в оперон и гена регулятора.
Мы остановились на том, что лактозный оперон похож на логический элемент AND. Но откуда у него берутся цифровые свойства? Ведь оба входных сигнала (концентрации цАМФ и лактозы), вообще-то, аналоговые. Попробуем нарисовать входные функции лактозного оперона.
Каждый вход лактозного репрессора состоит из двух этапов молекулярного узнавания. Лактоза связывается с лактозным репрессором, а репрессор — с ДНК. В клетке находится несколько десятков молекул репрессора, каждая из которых, пока нет лактозы, может связываться с началом лактозного оперона. Когда она там связана, РНК-полимераза не может начать работу. Из-за теплового движения молекулы репрессора то и дело отваливаются от ДНК и присоединяются обратно. Если репрессора нет совсем, оперон работает на полную мощность (сейчас для простоты считаем, что катаболический активатор есть в избытке). При малых концентрациях репрессора он снижает активность оперона практически линейно. Но дальше эффект каждой новой порции репрессора все меньше и меньше, и в целом график зависимости близок к гиперболе y = 1 / (x + 1)
Связывание репрессора с ДНК можно считать обратимой химической реакцией. Положение равновесия в обратимых химических реакциях описываются законом действующих масс. Что это такое?
Например, у нас есть реакция, в которой из веществ А и В образуется вещество С. Реакция обратима, то есть С может распадаться обратно на А и В в тех же самых условиях:
В колбе, где идет эта реакция, установится какое-то равновесие между прямым и обратным направлением реакции. В системе будут все три вещества А, В и С в каких-то концентрациях (в физической химии концентрацию вещества принято обозначать его символом в квадратных скобках, [A], [B] и [C] в нашем примере). Закон действующих масс связывает равновесные концентрации веществ между собой: произведение концентраций продуктов, деленое на произведение концентраций исходных веществ, постоянно.
Буквой К здесь обозначена константа равновесия реакции. Она зависит только от химической природы реакции (т. е. какие вещества реагируют) и от температуры. Концентрации веществ, давление, катализаторы и прочие факторы не влияют на константу равновесия.
Например, если мы добавим в колбу побольше исходного вещества А, то равновесие сдвинется в сторону вещества С, его концентрация возрастет, а концентрация вещества В (которого мы не добавляли) при этом упадет. Однако отношение [C] к ([A] * [B]) останется постоянным.
Итак, у нас есть обратимая реакция связывания активной формы репрессора с опероном: RepA + OpA Op (связывание репрессора превращает активную форму оперона в неактивную). Она имеет свою константу равновесия К
Из второго уравнения выражаем [Op] через [OpA] и подставляем в первое уравнение:
К
После преобразований получаем:
[OpA] = 1 / (K
То есть функция имеет вид y = 1 / (x + 1)
Молекулы репрессора плавают в клетке и связываются с ДНК только пачками по четыре. Чтобы активная форма репрессора превратилась в неактивную, она должна связаться с четырьмя молекулами лактозы — одной или двух недостаточно, надо по одной молекуле лактозы на каждую белковую субъединицу счетверенного репресора. Пока лактозы в клетке мало, ее молекулы связываются с репрессором по одной-две и он остается активным. Но после некоторой пороговой концентрации лактозы большинство молекул репрессора связывают по четыре молекулы лактозы и переходят в неактивную форму. График активности репрессора в зависимости от количества лактозы поэтому имеет S-образную форму и описывается функцией вида y = 1 / (1 + x^4).
Теперь найдем зависимость [RepA] от концентрации лактозы [Lac]. Она так же выводится из закона действующих масс, с одной тонкостью: в связывании с репрессором участвуют четыре одинаковых молекулы лактозы.
RepA + 4 Lac Rep
В уравнении для константы равновесия в этом случае появляются степени:
К
Опять мы используем тот факт, что суммарная концентрация обоих форм репрессора постоянна:
[Rep] + [RepA] = n, где n ≈ 50
И получаем, что [RepA] = n / (К
Теперь, чтобы получить входную характеристику оперона по лактозе, надо подставить вторую функцию в первую. Она будет с точностью до коэффициентов иметь такой вид:
y = 1 / (1 + (1 / (1 + x^4)))
Ее график тоже S-образно изгибается. Получается, что лактозный оперон не реагирует на малые концентрации лактозы. При достижении пороговой концентрации появляются неактивные молекулы репрессора, связавшие по четыре лактозы, и оперон довольно резко включается.
Второй регуляторный вход лактозного оперона тоже состоит из двух шагов молекулярного узнавания: цАМФ связывается с активатором, а активатор — с ДНК. Отличие от лактозного входа в том, что здесь связывание молекул (активатора или цАМФ) повышает активность того, с чем они связались (оперона или активатора). То есть графики будут выходить из нуля и приближаться к горизонтальной прямой где-то выше оси Х.
Поскольку связывание цАМФ включает активатор, активная форма будет продуктом реакции, а не исходным веществом:
2 цАМФ + Akt AktA
Константа равновесия K <Ц-А>= [AktA] / ([Akt] * [цАМФ]^2)
Опять мы пользуемся тем, что сумма концентраций [AktA] и [Akt] постоянна, так как в клетке есть около 100 молекул активатора, переходящих между активной и неактивной формой. Выражая [Akt] через [AktA], получаем:
K <Ц-А>= [AktA] / ((100 — [AktA]) * [цАМФ]^2)
[AktA] = K <Ц-А>* [цАМФ]^2 / (1 + K <Ц-А>* [цАМФ]^2)
То есть зависимость [AktA] от [цАМФ] имеет вид y = x^2 / (1 + x^2)
Из-за связывания двух молекул цАМФ график выглядит не как гипербола, а как S-образная кривая. Хотя пороговый эффект на ней заметен слабее, чем на графике для лактозы, потому что степень не четвертая, а только вторая.
Зависимость активности оперона от количества активатора имеет другой нюанс. Если связывание репрессора подавляет оперон почти полностью (где-то до 0,1%, что ниже погрешности биохимических измерений), то отсутствие активатора подавляет активность только до 5%. Поэтому график будет выходить не из нуля, а из точки (0; 0,05):
y = 0,05 + (0,95 * x / (1 + x))
Активность = [OpA] + 0,05 * [Op]
Зависимость [OpA] от [AktA] мы можем легко вывести по аналогии с предыдущими тремя. Активатор связывается по одной штуке, значит, [AktA] будет входить в формулу в первой степени. Он активирует оперон, значит, [AktA] будет и в числителе, и знаменателе:
[OpA] = K <А-O>* [AktA] / (K <А-O>* [AktA] + 1)
То есть функция имеет вид y = x / (1 + x)
И это только вклад оперона, связанного с активатором! Чтобы учесть вклад оперона без активатора, надо добавить слагаемое (1 — эта дробь) * 0,05. К счастью, оба слагаемых имеют общий знаменатель, поэтому функция усложняется совсем немного:
y = 0,05 + (0,95 * x / (1 + x))
Осталось получить зависимость активности оперона от концентрации цАМФ. Для этого надо подставить первую формулу (для концентрации активного CAP в зависимости от концентрации цАМФ) во вторую формулу (для активности оперона от концентрации активного САР)…
… И получить четырехэтажную дробь:
У нас получилась самая сложная функция из всех в этой статье. Впрочем, ее график выглядит похоже на график более простой зависимости активного CAP от концентрации цАМФ, y = x^2 / (1 + x^2):
Тоже S-образная кривая с некоторым пороговым эффектом, которая медленно приближается к горизонтали y = 1. Только начинается не из нуля, а из точки (0; 0,05).
Осталось понять, как взаимодействуют два входа. В случае лактозного оперона ответ простой — никак. Лактозный репрессор и катаболический активатор не влияют на связывание друг друга с ДНК. Поэтому связывание этих двух белков можно считать независимыми событиями. Полная активность оперона достигается, когда и активатор связан, и репрессор не связан. Вероятность такого совпадения равна произведению вероятностей каждого из них по отдельности. Значит, чтобы получить функцию активности лактозного оперона от двух переменных ([Lac]) и ([цАМФ]), надо просто перемножить функции от каждой этой переменной:
Активность = (1 / (1 + (1 / (1 + [Lac]^4)))) * (0,05 + 0,095 * ([цАМФ]^2 / (1 + [цАМФ]^2)) / (1 + ([цАМФ]^2 / (1 + [цАМФ]^2))))
Трехмерный график этой функции
похож на плато, обрывающееся ущельями по двум сторонам. Правое ущелье (малые концентрации лактозы) более глубокое и плоскодонное, чем левое (малые концентрации цАМФ).
В реальной жизни кишечной палочки вокруг нее лактозы либо нет совсем (чаще всего), либо ее концентрация выше пороговой и лактозный репрессор практически не мешает работе оперона (когда хозяин съел что-то молочное). Концентрация цАМФ — это внутренний сигнал, который вырабатывается самой клеткой. Она тоже бывает либо слишком малой для включения лактозного оперона (когда есть глюкоза или крахмал), либо достаточной для включения на 95% и более (если ничего вкуснее лактозы нет). То есть почти всегда лактозный оперон находится в условиях либо плато на этом графике, либо в одном из ущелий.
Теперь отвлечемся от лактозного оперона и посмотрим шире, на что способны логические элементы на такой химической базе. Практически везде в биохимических сигнальных системах мы встречаем молекулярное узнавание, которое описывается функциями вида y = x^n / (1 + x^n) для активаторов и y = 1 / (1 + x^n) для репрессоров. Степень n в этих функциях показывает количество связываемых однотипных молекул для получения эффекта, и бывает обычно равна 1, 2 или 4 (чаще всего 2). Можно получить более сложные логические ключи, добавляя в начало оперона больше сайтов связывания регуляторных белков. Этих белков может быть больше двух типов. Эффекты разных белков на посадку РНК-полимеразы могут складываться (OR) или перемножаться (AND). Но в целом мы будем иметь дело с какой-то комбинацией (сложением или перемножением) базовых функций y = x^n / (1 + x^n) и y = 1 / (1 + x^n) с разными степенями и коэффициентами.
Некоторые интересные функции получаются очень просто. Например, один и тот же регуляторный белок может связываться (в спаренной форме) с двумя участками промотора. В одном участке он действует как репрессор, а в другом — как активатор. Для работы оперона надо, чтобы сайт репрессора был пуст, а сайт активатора — занят. Получается функция с четким максимумом:
y = (1/ (1 + x^2)) * (x^2 / (1 + x^2))
Без этого белка оперон неактивен, потому что пуст сайт активатора, а при высоких концентрациях он неактивен, потому что занят сайт репрессора. Максимальная активность будет, когда и сайт активатора, и сайт репрессора заняты с вероятностью 50%.
Еще можно сделать два сайта связывания активатора, в одном активатор будет сильно связываться с ДНК, но слабо активировать ген, в другом — слабо прикрепляться к ДНК, но сильно активировать ген. В этом случае получается функция вроде y = x^4 / (1 + x^4) + 0.3 * (10x)^4 / (1 + (10x)^4), со ступенчатым графиком:
Математики, ау! Можете рассказать в комментариях, что еще можно и что нельзя получить при сложении и перемножении этих базовых функций y = x^n / (1 + x^n) и y = 1 / (1 + x^n)?
В следующей части займемся схемами из таких логических элементов.
Читайте также:
