
Кишечная палочка имеет жгутики для передвижения
ПОВЕДЕНИЕ БАКТЕРИЙ (ГРОМОВ Б.В. , 1997), БИОЛОГИЯ
Описан механизм работы "мотора", приводящего в движение бактериальные жгутики, охарактеризованы общие закономерности движения бактерий, природа хемотаксисов, вызывающих движение в определенном направлении.
Санкт-Петербургский государственный университет
Некоторые люди склонны считать, что способность к определенному поведению характерна только для человека, другие считают, что поведение присуще только животным, снабженным центральной нервной системой. Однако обсуждение особенностей поведения бактерий для специалистов представляется совершенно естественным несмотря на то, что бактерии - это самые примитивные организмы, имеющие клеточное строение, но лишенные клеточных органелл и цитоскелета.
Не существует общепринятого определения термина "поведение", различные его трактовки подробно обсуждаются в книге М.Г. Гаазе-Рапопорта и Д.А. Поспелова "От амебы до робота: модели поведения" (М.: Наука, 1987. 285 с.). Авторы приходят к выводу, что в наиболее общем понимании поведение "всегда представляется в виде процесса, в рамках которого реализуется взаимодействие организма с окружающей его средой. Поведение в самом общем виде - это формируемый организмом отклик на сигналы, поступившие к нему от окружающей среды" (с. 10). Следует все же добавить, что под поведением обычно понимают реакции, проявляющиеся в разного рода движениях. Под поведением бактерий понимают способность некоторых из них к направленному передвижению в соответствии с сигналами, поступающими из окружающей среды.
Около половины известных видов бактерий способны двигаться, причем существуют несколько типов движения бактерий. Мы будем говорить только о бактериях, способных плавать за счет работы жгутиков, в основном о кишечной палочке Escherichia coli, палочковидной бактерии, обитающей в кишечнике человека и животных и являющейся в то же время модельным объектом во многих биологических исследованиях.
Бактериальный жгутик - замечательное образование. Он состоит из спиральной нити, крюка и базальной структуры (рис. 1). Нить жгутика представляет собой жесткий полый цилиндр, образованный белковыми молекулами, уложенными в плотную спираль. Диаметр цилиндра около 120 нм. По длине нити белковые молекулы образуют 11 рядов. В процессе роста нити белковые молекулы, синтезированные внутри клетки, проходят через полость цилиндра и пристраиваются в спираль на ее конце. На конце жгутика имеется белковая шапочка или крышечка, закрывающая отверстие цилиндра и препятствующая выходу молекул белка в окружающую среду. Длина нити жгутика может достигать нескольких микрометров. Нить жгутика через белковые цилиндрики-соединители прикреплена к крюку. Крюк - относительно короткий цилиндр, образованный другим белком. Крюк обеспечивает мягкое соединение нити с палочковидной осью базальной структуры. Эта ось пронизывает клеточную стенку бактерии, на нее насажены четыре кольца. Три внешних кольца служат для фиксации оси в стенке бактерии, тогда как четвертое, встроенное в цитоплазматическую мембрану клетки представляет собой как бы вращающийся ротор электрического мотора, которым является базальная структура бактериального жгутика.
Любая бактерия может существовать, только если на ее цитоплазматической мембране функционирует протондвижущая сила, заставляющая протоны заходить внутрь клетки. Протондвижущая сила обеспечивается разностью концентраций протонов на внешней и внутренней сторонах мембраны (на внешней стороне их больше) и наличием более отрицательного заряда на внутренней стороне мембраны, протоны же, как известно, несут положительный заряд. У бактерий, живущих в средах с высокой концентрацией соли, вместо протонов могут быть использованы катионы натрия. Протондвижущая сила обеспечивает многие жизненно важные для бактерий процессы обмена клетки с окружающей средой, кроме того, она используется при работе жгутика. По существующей гипотезе внутреннее, встроенное в цитоплазматическую мембрану кольцо базальной структуры, то есть вращающийся ротор, окружено мембранными белками, имеющими определенным образом пространственно организованные отрицательные заряды - это статор мотора. Протондвижущая сила заставляет протоны проходить через базальную структуру внутрь клетки, при этом в какой-то момент они задерживаются на определенных участках ротора, придавая им положительный заряд, затем протоны уходят внутрь клетки. Заряженные участки расположены таким образом, что возникает сила притяжения между заряженными участками ротора и статора, кольцо начинает вращаться. Установлено, что для полного оборота кольца через базальную структуру должно пройти 500-1000 протонов. Вращение кольца через жестко связанную с ним ось и крюк передается нити жгутика, которая функционирует как пропеллер или корабельный винт. Кольцо совершает около 300 оборотов в секунду. Нить жгутика представляет собой жесткую спираль, закрученную против хода часовой стрелки. Если кольцо также вращается против часовой стрелки, нить как бы ввинчивается в окружающий раствор. На работу жгутика бактерия тратит около 0,1% всей расходуемой ею энергии.
Число жгутиков у разных бактерий может быть различным: у кишечной палочки 6-7 жгутиков, расположенных в разных местах клеточной поверхности. Если жгутики вращаются против часовой стрелки, их нити накладываются одна на другую и образуется единая спираль (рис. 2), лучше обеспечивающая движение бактерии. При вращении жгутика клетка бактерии, естественно, тоже вращается в противоположном направлении, но число оборотов невелико, поскольку клетка намного массивнее жгутика. По всей видимости, поскольку в процессе движения клетка все время вращается, она неспособна воспринимать направление силы тяжести, то есть не обладает способностью к гравитаксису. Во всяком случае пока нет никаких свидетельств наличия у бактерий такой способности.
Однако пришло время обратить внимание на то, что из-за мелких размеров клеток в силу законов физики бактерии в водной среде находятся в условиях, совсем не похожих на те, в которые попадает пловец. Важной гидродинамической характеристикой плывущего предмета является число Рейнольдса, представляющее собой отношение сил инерции к силам вязкости. С уменьшением размеров объекта это число уменьшается, поэтому водная среда является для бактерий вязкой, каковой для нас была бы патока. Бактерия плывет до тех пор, пока работает винт, вклад инерции исключительно мал. Кроме того, бактерии, даже мертвые, находящиеся в водной среде, перемещаются в результате так называемого броуновского движения, бактериальная клетка все время подвержена ударам окружающих молекул, находящихся в тепловом движении. Удары, наносимые с разных сторон, бросают бактерию из стороны в сторону.
Подсчитано, что плыть целенаправленно в одном направлении бактерия может не более 3 с, затем удары окружающих молекул разворачивают ее в случайном направлении. Видимо, в связи с этим бактерии выработали собственный механизм изменения направления движения. Когда нить жгутика вращается против хода часовой стрелки, бактерия движется приблизительно по прямой, эта стадия движения определяется как пробег. Скорость перемещения в период пробега у разных бактерий обычно бывает в пределах 20-80 мкм/с. Кишечная палочка плывет со скоростью около 30 мкм/с, нетрудно рассчитать, что, если бы даже она могла плыть все время прямо, ей удалось бы за 1 ч проделать путь всего лишь в 10 см. Правда, несколько иная оценка получится, если соотнести скорость передвижения с длиной тела организма. Длина молодой кишечной палочки около 1,7 мкм, многие люди имеют рост 170 см, то есть человек в миллион раз длиннее. Если бы он двигался относительно длины своего тела с той же скоростью, что и кишечная палочка, то его скорость составляла около 100 км/ч. Пробег бактерии, однако, длится около одной секунды, после чего происходит переключение жгутикового мотора, который начинает вращаться по ходу часовой стрелки, что приводит к остановке и развороту бактерии в случайном направлении. Переключение мотора занимает менее 1 мс. У кишечной палочки при этом единая жгутиковая спираль распадается, жгутики разлетаются в разные стороны и бактерия совершает "кувырок" (см. рис. 2). Мотор вращается по ходу часовой стрелки всего около 0,1 с, после чего снова переходит к вращению против часовой стрелки, и бактерия опять плывет по прямой, но уже в другом, случайном направлении. Длительность пробега или частота кувырков, что одно и то же, в некоторых пределах постоянны и зависят от организма и условий среды. Замечено, что этот показатель, кроме того, характеризует конкретную бактерию, придавая ей индивидуальность. После деления две дочерние клетки приобретут новые собственные характеристики, это, разумеется, ненаследуемые индивидуальные свойства организмов.
Пока окружающая среда остается неизменной, бактерии плавают беспорядочно, такое передвижение не имеет смысла, так же как заблудившийся в лесу человек, не имеющий ориентиров, проходит многие километры, не удаляясь значительно от исходной точки своего пути.
Установлено, что само по себе плавание не дает бактерии никаких преимуществ. Однако совершенно гомогенной окружающая среда бывает редко даже в лабораторных условиях. Если среда неоднородна, бактерии в некоторых случаях могут определять эту неоднородность, тогда движение становится целенаправленным - это так называемые таксисы, которые можно рассматривать как элементарные поведенческие реакции.
Впервые на это явление еще в 90-х годах прошлого столетия обратили внимание немецкие ученые. Теодор Энгельманн обнаружил, что в водных микроскопических препаратах можно видеть, как некоторые бактерии скапливаются вокруг пузырьков воздуха, попавших под покровное стекло, тогда как другие от этих пузырьков убегают. В первом случае бактерии нуждались в молекулярном кислороде, а во втором он был для них вреден. Это явление получило название аэротаксиса. Вильгельм Пфеффер помещал в воду, содержащую бактерии, капилляр, наполненный раствором различных веществ. Иногда бактерии скапливались у окончания капилляра и даже набивались внутрь. Это явление определяют как хемотаксис. В опытах Пфеффера бактерий привлекали сахара или пептон - вещества, используемые ими в качестве пищи. Однако уже Пфеффер показал относительность целесообразности хемотаксиса: он помещал в капилляр с пептоном сулему - яд, который бактерии не замечали и, набиваясь в капилляр, умирали. После описанных наблюдений микробиологи на долгие годы потеряли интерес к изучению поведения бактерий, вероятно, потому, что в то время не было адекватных методов изучения этого явления.
Только в 60-х годах нашего столетия американский ученый Юлиус Адлер в университете штата Висконсин продолжил исследования хемотаксиса бактерий, уже, конечно, на совсем другом методическом уровне. Сразу же были обнаружены интереснейшие явления, и исследования поведения бактерий начали развиваться лавинообразно. В настоящее время в этой области накоплен огромный материал, объясняющий наблюдаемые явления с позиций биофизики, молекулярной биологии и молекулярной генетики.
Бактерии способны реагировать не на любые соединения, но только на определенные и различные для разных бактерий. Такие вещества называют хемоэффекторами. Среди эффекторов есть вещества, привлекающие бактерий, - аттрактанты, и вещества, их отпугивающие, - репелленты. В поверхностных структурах бактериальной клетки есть специальные белковые молекулы - рецепторы, специфически соединяющиеся с определенным эффектором, при этом молекула эффектора не изменяется, а в молекуле рецептора происходят конформационные и другие изменения. У большинства бактерий есть рецепторы, воспринимающие молекулярный кислород, протоны, у многих есть рецепторы аминокислот, сахаров. Кишечная палочка имеет около 20 различных типов рецепторов, всего клетка кишечной палочки содержит около 25 тыс. молекул рецепторов. Рецепторы расположены не равномерно по всей поверхности клетки, а сконцентрированы на одном из полюсов. Состояние рецептора отражает внеклеточную концентрацию соответствующего эффектора. У кишечной палочки в качестве важных компонентов сенсорной системы функционируют четыре так называемых метилакцепторных белка, которые являются рецепторами или получают сигналы от других рецепторов. Эти белки способны присоединять к заряженным глютаминным остаткам метильные группы или отдавать их за счет активности соответствующих ферментов, причем уровень метилирования белка соответствует концентрации эффектора. Следует заметить, что у разных бактерий в этом отношении обнаруживаются различия, а в некоторых таксисах метилакцепторные белки участия не принимают, например в таксисе к молекулярному кислороду у кишечной палочки.
Клетки бактерий, находящиеся в течение некоторого времени в средах с разной концентрацией сахара, ведут себя совершенно одинаково, но уровень метилирования соответствующего белка у них различен. Бактерии способны воспринимать изменение концентрации эффектора во времени, поскольку, если концентрация изменяется, она уже не соответствует уровню метилирования метилакцепторного белка. В этом случае сигнал поступает на определенные цитоплазматические белки, изменяя уровень их фосфорилирования, а эти белки передают сигнал уже на жгутиковый мотор, в результате чего изменяется длина пробега. При увеличении концентрации аттрактанта длительность пробега возрастает (а частота кувырков соответственно уменьшается), при увеличении концентрации репеллента длительность пробега уменьшается. При снижении концентраций эффекторов наблюдается обратный эффект. Это, очевидно, означает, что бактерии имеют элементарную память и "помнят", какой была концентрация эффектора в предыдущий момент. Эта память у них, правда, "девичья", обычно через 2-3 мин в результате процесса адаптации уровень метилирования белка приходит в соответствие с новой концентрацией эффектора, и длина пробега возвращается к норме. Однако, если в среде создается градиент концентрации эффектора, в результате многочисленных изменений длины пробега в зависимости от направления движения клетки бактерии постепенно приближаются к источнику аттрактанта или удаляются от репеллента (рис. 3).
Как и следовало ожидать, в качестве аттрактантов обычно выступают вещества, для бактерий полезные, а в качестве репеллентов вредные. Дело, однако, обстоит не столь просто. Для кишечной палочки эффекторами (аттрактантами или репеллентами) служат только некоторые из аминокислот, причем нет связи с возможностью их использования бактерией. Для той же бактерии ацетат является репеллентом, хотя даже может быть использован в качестве пищевого субстрата и т.п. Поэтому существует мнение, что в действительности эффекторы воспринимаются бактериями как сигналы об определенной экологической обстановке. Наличие какой-либо аминокислоты, например, может свидетельствовать о присутствии разлагающегося белка. Ацетат и другие органические кислоты образуются в процессах сбраживания сахаров и полисахаридов и т.п.
Приспособительное значение хемотаксиса не вызывает сомнений, а в некоторых случаях экспериментально доказано. Например, формы холерного вибриона с нарушенным хемотаксисом оказываются менее болезнетворными (вирулентными), чем исходные. За счет хемотаксиса к корешкам растений приближаются симбиотические или паразитические бактерии, корневые выделения иногда воспринимаются ими с расстояния до 10 см (огромное для бактерии расстояние).
Кроме хемотаксиса бактерии могут проявлять и другие поведенческие реакции. Это прежде всего фототаксис, характерный для бактерий, использующих свет в качестве источника энергии. Для некоторых патогенных (болезнетворных) бактерий большое значение имеет способность к вискозитаксису - бактерии стремятся в среду с большей вязкостью, существует и термотаксис - движение в сторону повышения или понижения температуры.
Особенно замечательна способность некоторых бактерий плыть вдоль линий магнитного поля - магнетотаксис. В клетках таких бактерий, называемых магнетобактериями, находятся кристаллики железосодержащих минералов (например, магнетита), ориентирующиеся вдоль линий магнитного поля как стрелка компаса. Железо составляет около 3% сухой массы магнетобактерий. Это водные бактерии, обитающие в пресных водоемах и море. Они плывут по линиям магнитного поля Земли, причем в северном полушарии к северному полюсу, а в южном полушарии к южному. Это может показаться странным, дело, однако, в том, что, плывя таким образом, бактерии углубляются в воду в результате того, что их магнитосомы ориентируются по результирующей вертикальной и горизонтальной составляющим магнитного поля. Чем ближе к северу, тем круче они уходят в воду и попадают на поверхность ила, где, очевидно, больше пищи. Кроме того, магнетобактерии лучше себя чувствуют при незначительном содержании молекулярного кислорода, что как раз и наблюдается в поверхностных слоях ила. Как было отмечено, бактерии, видимо, неспособны к гравитаксису и только использование линий магнитного поля Земли дает им возможность различать верх и низ.
Результаты изучения движения и поведения бактерий представляют общенаучный интерес. В самом деле, бактериальная клетка снабжена электрическим мотором, да еще работающим на протонах. У высших организмов ничего подобного не обнаружено. Механизмы рецепции сигналов в хемотаксисе, их передачи и реализации сейчас хорошо изучены. Очевидно, что они оригинальны и значительно отличаются от механизмов, используемых в сенсорных системах организмов с эукариотической клеткой. Видимо, здесь эволюция бактерий шла своим путем.
1. Каппуччинелли П. Подвижность живых клеток. М.: Мир, 1982. 125 с.
2. Moens S., Vanderleyden J. Functions of Bacterial Flagella // Crit. Rev. Microbiol. 1996. Vol. 22. P. 67-100.
Борис Васильевич Громов, доктор биологических наук, профессор, член-корреспондент РАН, зав. кафедрой микробиологии Санкт-Петербургского государственного университета. Область научных интересов - общая и экологическая микробиология, главным образом исследования фототрофных микроорганизмов. Автор 220 печатных работ в отечественных и международных изданиях, в том числе двух монографий и учебных руководств.
По существующей гипотезе, внутреннее, встроенное в цитоплазматическую мембрану кольцо базальной структуры - вращающийся ротор, окружено мембранными белками, имеющими определенным образом пространственно организованные отрицательные заряды - статор мотора. Протонодвижущая сила заставляет протоны проходить через базальную структуру внутрь клетки, при этом в какой-то момент они задерживаются в определенных участках ротора, придавая им положительный заряд, затем протоны уходят внутрь клетки. Заряженные участки расположены таким образом, что возникает сила притяжения между заряженными участками ротора и статора, кольцо начинает вращаться. установлено, что для полного оборота кольца через базальную структуру должно пройти 500-1000 протонов. Кольцо совершает около 300 оборотов в секунду. Нить жгутика закручена против ч.с. Если кольцо вращается против ч.с. нить ввинчивается в окружающий раствор. На работу жгутика бактерия тратит около 0,1% расходуемой энергии. У кишечной палочки 6-7 жгутиков. Бактерии не воспринимают направление силы тяжести из-за вращения клетки.
Важной гидродинамической характеристикой плывущего предмета является число Рейнольдса представляющее собой отношения сил инерции
к силам вязкости. С уменьшением размеров объекта это число уменьшается, поэтому водная среда является для бактерий вязкой, каковой для нас была бы патока. Подсчитанно, что плыть целенаправленно в одном направлении бактерия может не более 3 с, затем удары окружающих молекул разворачивают ее в случайном направлении.
Когда нить жгутика вращается против ч.с. (рис.2) бактерия движется по прямой - пробег. Скорость движения во время пробега для разных бактерий 20-80 мкм/с. Для кишечной палочки 30 мкм/с т.е. 10 см/ч. Пробег длится около секунды, после чего происходит переключение жгутикового мотора, который начинает вращаться по ходу ч.с. что приводит к остановке и развороту бактерии в случайном направлении. Переключение занимает менее 1 мс. У кишечной палочки при этом единая жгутиковая спираль распадается и бактерия совершает кувырок. Мотор вращается по ходу ч.с. около 0.1 с после чего снова переходит к вращению против ч.с.
Кишечная палочка имеет около 20 различных типов рецепторов, всего клетка кишечной палочки содержит около 25 тыс. молекул рецепторов. Рецепторы сконцентрированы на одном из полюсов.
У кишечной палочки в качестве важных компонентов сенсорной системы функционируют четыре так называемых метилакцепторных белка, которые являются рецепторами или получают сигналы от других рецепторов . Эти белки способны присоединять к заряженным глютаминным остаткам метильные группы или отдавать их за счет активности соответствующих ферментов, причем уровень метилирования белка соответствует концентрации эффектора. Не во всех таксисах так происходит.
Бактерии способны воспринимать изменение концентрации эффектора во времени, поскольку, если концентрация изменяется, она уже не соответствует уровню метилирования метилакцепторного белка. В этом случае сигнал поступает на определенные цитоплазматические белки, изменяя уровень их фосфорилирования, а эти белки передают сигнал уже на жгутиковый мотор, в результате чего изменяется длина пробега.
При увеличении концентрации аттрактанта длительность пробега возрастает ( а частота кувырков уменьшается), при увеличении концентрации реппелента длительность пробега уменьшается. При снижении концентрации эффекторов наблюдается обратный эффект. Это означает, что бактерии имеют элементарную память.
Обычно через 2-3 мин в результате процесса адаптации уровень метилирования белка приходит в соответствие с новой концентрацией эффектора и длина пробега возвращается к норме. Однако, если в среде создается градиент концентрации эффектора, в результате многочисленных изменений длины
пробега в зависимости от направления движения клетки бактерии постепенно приближаются к источнику аттрактанта или удаляются от реппелента (рис. 3)
Магнетотаксис - способность двигаться вдоль линий магнитного поля. В клетках таких бактерий находятся кристаллики железосодержащих минералов (напр. магнетит). Железо составляет около 3% сухой массы магнетобактерий. Это водные бактерии обитающие в пресных водоемах и море. В северном полушарии они плывут к северному полюсу и наоборот. Бактерии углубляются в воду т.к. магнетосомы ориентируются по результирующей вертикальной и горизонтальной составляющим магнитного поля. Чем ближе север тем круче они уходят в глубину в ил, где много пищи и мало кислорода.
Одним из крупнейших достижений человека на заре его развития было создание постоянного источника продуктов питания путем одомашнивания диких животных и возделывания растений. Создание разнообразных пород животных и сортов растений стало возможным благодаря разработке принципов искусственного отбора.
Окультуривание диких растений происходило независимо в различных географических областях и основывалась на местной флоре. Распространение культурных растений за пределы центра происхождения происходило как естественным путем, так и вследствие миграций населения. Так возникли первичные и вторичные центры происхождения и разнообразия культурных растений. Открытие центров происхождения культурных растений принадлежит выдающемуся русскому ученому Н.В.Вавилову, собравшему огромный семенной материал для использования в селекционной работе.
Было выделено восемь центров происхождения культурных растений:
1. Средиземноморский (спаржа, маслины. капуста, лук, клевер, мак, свекла, морковь)
2. Переднеазитский (инжир, миндаль, виноград, гранат, люцерна, рожь, дыня, роза)
3. Среднеазиатский (нут, абрикос, горох, груша, чечевица, лен, чеснок, мягкая пшеница)
4. Индо-Малайский (цитрусовые, хлебное дерево, огурец, манго, черный перец, кокосовая пальма, банан, баклажан)
5. Китайский (просо, редька, вишня, яблоко, гречиха, слива, соя, хурма)
6. Центрально-американский (тыква, фасоль, какао, авокадо, махорка, кукуруза, батат, хлопчатник)
7. Южноамериканский (табак, ананас, томат, картофель)
8. Абиссинский центр (банан, кофе, сорго, твердая пшеница)
В настоящее время выделяют 12 первичных центров происхождения культурных растений:
1. Китайско-Японский
2. Индонезийско-Индокитайский
3. Австралийский
4. Индостанский
5. Среднеазиатский
6. Переднеазиатский
7. Средиземноморский
8. Африканский
9. Европейско-Сибирский
10. Центральноамериканский
11. Южно-Американский
12. Северо-Американский
Циклы могут быть эндогенной природы и вызванные внешними периодическими изменениями
Циркадные ритмы привышают время генерации клетки или особи, т.е. лунные ритмы не встречаются у организмов период генерации которых меньше месяца, циркадные колебатели известны для тех клеток, время генерации которых превышает 24 часа. Свет, температура, длина светового дня - колебатели циклов.
Время генерации эукариотических клеток близко к 24 часам. В клетках Acetobularia лишенных ядра циркадианные ритмы фотосинтеза сохранялись 30 суток - цитоплазмы достаточно для поддержания активности колебателя.
Медоносные пчелы, найдя источник пищи, напр. в 3 ч дня, на следующий день прилетают к тому же самому месту точнов то же самое время - память времени. Если крыс, которые обычно едят в начале ночи, кормить только утром, то подъем активности ферментов в тонком кишечнике смещается на раннее утро и сохраняется в течении нескольких циклов после предоставления животному свободного доступа к пище. Благодаря изменении
длины дня защитные стратегии (зимняя спячка, эстивация, покой, диапауза) у животных включаются раньше наступления неблагоприятного времени. Изменяемое время регистрируется вплоть до минут и механизм такого измерения не выяснен.
У млекопитающих колебатель ритма расположен около зрительной хиазми, где к нему пододят нервные волокна, идущие от сетчатки.
У тараканов циркадианные осцилляторы находятся в левой и правой зрительной доле. У воробъя - эпифиз. У бабочек - сатурний циркадианный римт выхода имаго исчезает после обезглавливания куколок, но восстанавливается, если пересадить куколкам мозго в гемоцель брюшка. У Apliysia оба глаза являются автономными колебателями. У животных и человека имеется несколько осцилляторов. Осцилляторы нервной природы.
У дневных птиц типичный суточный профиль активности включает один пик после рассвета и второй перед закатом (пение, крики, кормление, ухаживание, спаривание, защита территории). Утренний пик больше - в это время происходит строительство гнезда.
Социальная синхронизация ритмов - крики и пение птиц разных особей служит времязадателем.
ПРИЛИВЫ
полусуточные приливы 12,4 ч преобладают на побережьях Атлантического океана, Северного моря и в нек районах Индийского океана. Амплитуда суточных пиков одинакова и меняется в течении синодического месяца (от новолуния до новолуния Т=29,53 дня). Максимальные приливы в сизигии - луна, земля и солнце на одной прямой - новолуние, минимальные приливы в квадратуре.
Суточные приливы - 24 ч
циркадный ритм Дрозофилы требует взаимодействия по крайней мере двух генов period (per) и timeless (tim) Их транскрипты накапливаются днем и уменьшаются ночью. Эти молекулярные ритмы результат негативной обратной связи, когда Per и Tim белки взаимодействуют проникая в ядро и ингибируют транскрипцию своих собственных генов. Свет быстро разрушает tim, транскрипция возобновляется.
23 декабря 2014
- 9206
- 7,4
- 4
- 1
Несмотря на кажущуюся примитивность, бактерии не так уж просто устроены!
Бактерии — крохотные существа, и многие считают, что они очень просто устроены. Конечно, каждая бактерия — это всего лишь одна клетка, у которой нет отдельных частей тела, вроде ног или рук, нет глаз и носа, нет даже клеточного ядра. Но каким-то образом бактерии выживают и умудряются процветать с такими, казалось бы, ограниченными размерами и возможностями, да к тому же находить целое множество оригинальных решений для облегчения собственной жизни. Например, чтобы двигаться — то есть влиять на свое положение в пространстве самостоятельно, а не дожидаясь попутного течения, — бактерии придумывают самые удивительные ухищрения. Конечно, вы уже наверняка слышали о жгутиках бактерий. А что такое твитчинг? И как можно сдвинуть себя с места с помощью сахаров? Давайте присмотримся к бактериям чуть ближе. И сразу обратим внимание на то, из чего бактерии собирают себе средство передвижения.
Белковая диета
Среди подвижных бактерий больше всего тех, кто для движения использует белковые молекулы. Как они их применяют? Многие бактерии синтезируют специальные белки, из которых собирают подвижную ниточку — жгутик (рис. 1). Жгутик состоит из трех частей — филамента (собственно нити), крюка и базального тела. Каждая из этих частей сложена из белков. У хорошо изученной бактерии — кишечной палочки — белки, образующие филамент, называют флагеллинами и обозначают буквами Flg, Fli, Flh (от латинского слова flagellum — жгутик). Флагеллины складываются в нить, которая с помощью крюка крепится к базальному телу. Базальное тело — это что-то вроде якоря, который прочно закреплен в клеточной оболочке и может свободно вращаться по часовой стрелке или против. У бактерий может быть один или несколько жгутиков.
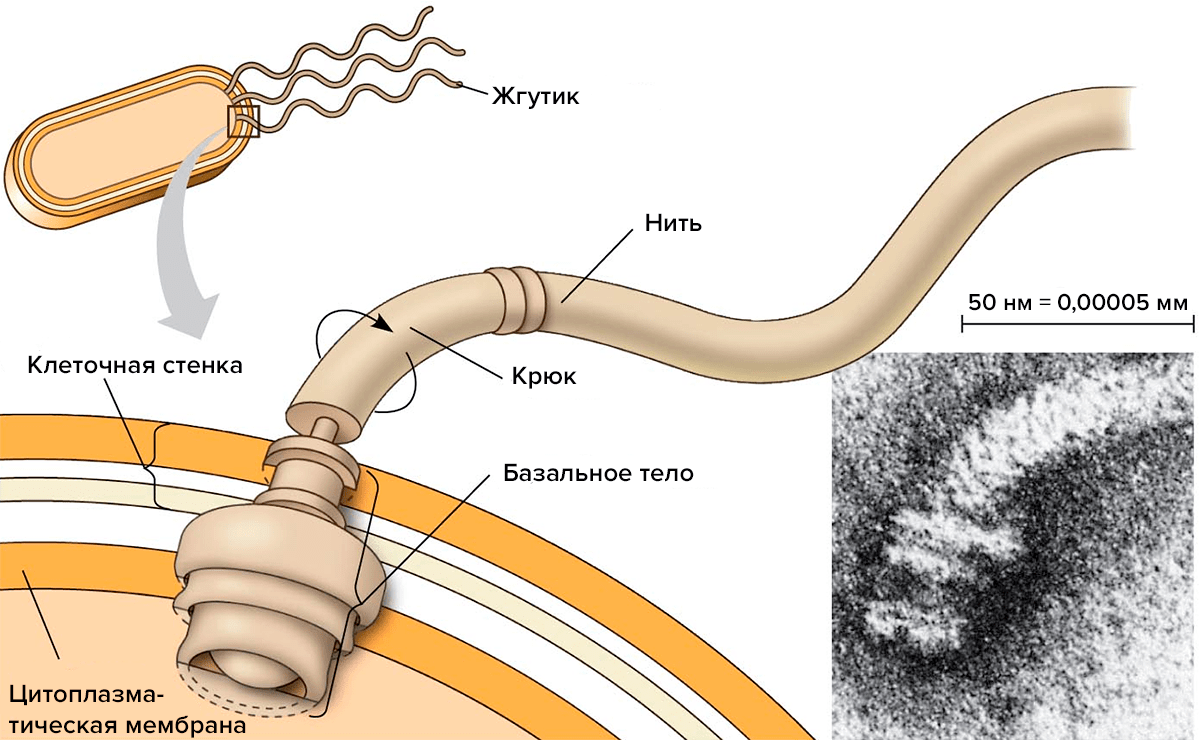
Рисунок 1. Строение жгутика.
Какие виды движения обеспечивает жгутик? Если бактерия находится в жидкой среде, то жгутик помогает ей плыть. Плавание — это самый быстрый способ передвижения. Причем, бактерия может неплохо управлять своим движением, меняя направление вращения базального тела: вращение базального тела по часовой стрелке толкает клетку в направлении от жгутика, а биения против часовой стрелки тянут клетку вслед за жгутиком.
А теперь представьте размахивание жгутиками на твердой поверхности, смоченной жидкостью. Бактерии будут не плыть, а расползаться в одной плоскости. Такое движение называется роением. Роение чаще бывает у бактерий, живущих в крупных колониях, — подвижные бактерии, находящиеся с краю, пытаются отодвинуться как можно дальше и основать свои собственные колонии.

Рисунок 2. Движение с помощью подтягивания к месту прикрепления пили.
Бактерии могут также создавать более короткие и просто устроенные нити, чем жгутики, — пили. Клетка может с помощью пили прикрепиться к чему-нибудь твердому, а потом подтянуться к месту крепления, разбирая эту нить, начиная от места крепления пили к клетке (рис. 2). Можно сказать, что клетка перемещается рывками. Подобный способ движения у одной клетки называется твитчинг (англ. twitch — дергать, тащить). А если так действует несколько скрепленных друг с другом бактерий, то они дружно скользят по твердой поверхности.
В оболочках клетки могут быть разнообразные белковые комплексы, например, обхватывающие клетку кольца из белков. Эти кольца крутятся, как гусеницы у гусеничного трактора, и помогают бактерии скользить по твердой поверхности. Такой способ подвижности есть у бактерии Flavobacterium johnsoniae.
У других бактерий есть белки, расположенные вдоль всей поверхности клетки. Эти белки создают продольные волны, и бактерия извивается и плывет в водной толще или скользит на твердом субстрате.
Очень многие микроорганизмы способны двигаться, но вот конкретный механизм или набор из нескольких механизмов у каждой специфичен. Поэтому, например, и говорят, что такое строение жгутика характерно для кишечной палочки, а у другой бактерии, тоже плывущей с помощью жгутика, все может быть устроено иначе — и ученые исследуют каждую бактерию по отдельности.
Как вы, наверное, заметили, пока что описанные способы движения позволяли бактерии плыть или скользить в зависимости от того, где она находится, — в жидкости или на твердой поверхности. Но скольжение может быть и единственным доступным способом передвижения.
Сахарный след
Многие бактерии выделяют наружу сахара. Смешиваясь с водой, сахара образуют слизь. Слизь облегчает движение клеток по твердой поверхности при использовании жгутиков.
Однако и сама слизь может быть источником движения. Представьте себе, что вы надуваете воздушный шарик. Внезапно шар вырывается из ваших рук и улетает под силой струи воздуха, резко выходящей из шарика. Подобным образом могут толкать себя и бактерии.
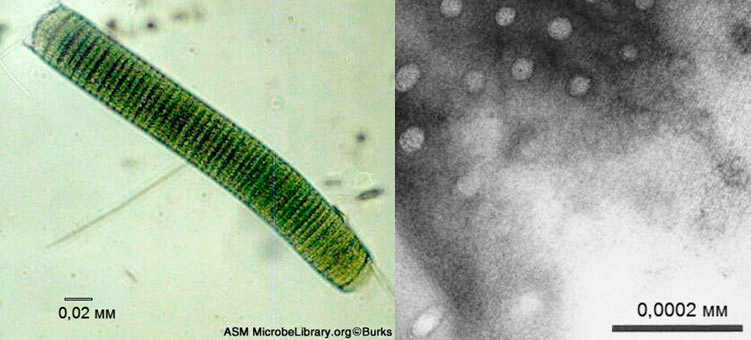
Рисунок 3. Oscillatoria princeps. Слева: Бактерии вида Oscillatoria princeps образуют нить. Справа: Большие поры на поверхности бактерии.
Выделение сахаров из специальных пор или биение жгутиков — это активные способы передвижения клетки. Бактерия взаимодействует с окружающей средой и активно отталкивается от воды или твердой поверхности. Но существует и пассивная подвижность, когда изменения внутри клетки приводят к ее перемещению благодаря внешним силам, например, току воды.
Газовые баллоны
Бактерии могут изменять свою плавучесть, накапливая внутри атмосферный воздух. Воздух все время диффундирует в толщу воды, а бактерии могут специально отбирать и накапливать молекулы разных газов в специальном баллоне, сложенном из белка. Таким образом клетка меняет свою плотность, начинает весить меньше и всплывает, выталкиваемая архимедовой силой. Если бактерии затем нужно погрузиться обратно, она может избавиться от воздуха или накопить внутри себя тяжелые сахара.
В какую сторону плыть?
Чтобы оказаться в самых подходящих для себя условиях, многие бактерии передвигаются не случайным образом, а целенаправленно, приближаясь к какому-нибудь приятному для себя объекту (например, еде или свету) или отплывая как можно дальше от неприятного (например, молекул, выделяемых другими бактериями). Такое целенаправленное движение называется таксисом. Чтобы распознавать сигналы из внешнего мира, бактерия синтезирует специальные белки — рецепторы, которые располагаются у нее на поверхности. Каждый вид рецепторов реагирует на свой стимул — молекулы еды, свет и так далее. Обнаружив свой стимул, рецептор передает сигнал о нем внутрь клетки.
Рисунок 4. Движение бактерии в направлении привлекающей ее пищи.
Если у клетки нет жгутика, то двигаться целенаправленно ей куда труднее. Но и тут можно что-то придумать. Например, газовые баллоны внутри клетки смещают бактерию вверх и вниз, то приближая ее к поверхности водоема, кислороду и свету, то погружая на дно.
Микроигра
Попробуйте расшифровать четыре слова, связанных с микробиологией, прослеживая движение бактерий к еде.
Каждая бактерия начинает двигаться от буквы, на которую указывает исходящая от бактерии стрелка. Затем бактерия меняет направление движения, согласно маленькой черной стрелочке у буквы. Если бактерия приближается к еде (красная точка), то она проплывает три клеточки (по горизонтали, вертикали или диагонали напрямую), снова поворачивает согласно направлению стрелки на этой клеточке и так далее. Если бактерия плывет в противоположную от еды сторону, она сдвигается только на одну клеточку. Если движение скорее нейтрально, то бактерия проплывает две клеточки.
Читайте также:
