Читайте также:
- А- как экономические отношения;
- Административно-правовые нормы и правоотношения
- Алексеев. Правовой статус личности — это правовое положение человека, отражающее его фактическое состояние во взаимоотношениях с обществом и государством.
- Б) сохраняют силу, кроме случаев, когда в законе установлено, что его действие распространяется на отношения, возникшие из ранее заключенных договоров;
- Билет № 4 Конституционно-правовые отношения: понятие, особенности, виды
- Билет №17. Экономико-математические модели в компьютерном управлении.
- Билет №27 Сетевые модели. Детерминированные, вероятностные модели.
- Билет №32. Критический путь в сетевой модели. Минимизация общей продолжительности проекта с учетом трудовых ресурсов.
- Блоковая модель документа. Содержимое блока, рамки, поля и отступы. Блочные и строчные теги как элементы блоковой модели. Свойства display, overflow.
- Бюджетные правоотношения и нормы бюджетного права.
|
Хищник – жертва.
Некоторые животные способны менять окраску в зависимости от цвета фона. Например, головоногие моллюски, некоторые ракообразные, рыбы, амфибии, рептилии. Экспериментально показано, что из всех рыб, выживших в водоёме, где охотился пингвин, 66% составили особи, окрашенные соответственно фону.
Взаимодействия хищников и их жертв очень динамичны и взаимообусловлены. Это особенно ярко выражается в проявлениях адаптивного поведения. Наблюдения за головастиками американской жабы Bufo americans показали, что в присутствии хищных личинок стрекоз они снижают активность, изменяется и их распределение в пространстве. Например, у коловраток Brahionus calchforus в присутствии хищной коловратки Asplanchna развиваются длинные боковые выросты панциря, которые препятствуют заглатыванию их. Образование этих шпор индуцируется веществом выделяемым хищником и уменьшением пищи.
Паразит – хозяин.
Паразитизм – это форма взаимоотношений двух различных видов организмов, носящих онтогенетический характер, при которой организмы одного вида (паразита, потребителя) живут за счёт питательных веществ или тканей организма другого вида (хозяина) в течение определённого времени. Паразитизм известен на всех уровнях организации живого, начиная с вирусов и бактерий и заканчивая высокоорганизованными животными.
Распространение паразитизма. Чем сложнее устроен организм, тем выше эффективность его гомеостазирующих систем и поэтому больше благоприятных возможностей он предоставляет в качестве среды обитания. Чем организм совершеннее, тем меньше становится для него потребность использовать благоприятные условия в другом организме. Это объясняет, почему явление паразитизма наиболее представлено среди микроорганизмов и примитивных многоклеточных. Число видов, ведущих паразитический образ жизни, резко падает у высокоорганизованных организмов. У высших животных встречается наиболее обильное разнообразие населения паразитов.
Формы паразитизма и взаимные адаптации паразитов и их хозяев.
· Эктопаразитизм (клещи, напр. таёжный клещ, собачий клещ; блохи);
· Эндопаразитизм (паразитические черви, напр. печёночный сосальщик, широкий лентец).
По степени тесноты связей паразита и хозяина выделяют 2 формы:
1. Облигатный – ведёт только паразитический образ жизни, не выживает без обязательной связи с хозяином (все черви паразиты, напр. Taeniarhynchus saginatus – бычий цепень). Характеризуется наиболее полным комплексом приспособлений к использованию организма хозяина как среды.
2. Факультативный – ведут свободный образ жизни и лишь при особых условиях переходят к паразитическому существованию. Например, Armillaria mellea – опёнок. Их связь исключительно трофическая. Морфофизиологические и биологические адаптации к паразитизму выражены слабо.
Адаптации парзитов: среда в виде живого и подвижного организма требует формирования способов фиксации в ней: зацепки, крючья, присоски, видоизменения формы тела. Эктопаразиты имеют сплющенное тело. У эндопаразитов утрачиваются органы, которые не нужны им в среде хозяина.
Математические модели.
В естественных условиях (в гетерогенной среде) возникает следующая временная и причинно-следственная цепь: размножение жертвы→размножение хищника→резкое сокращение численности жертвы→падение численности хищника→размножение жертвы и т.д.
Волны жизни хищника и жертвы следуют друг за другом с постоянным сдвигом по фазе, и в среднем численность как хищника так и жертвы остаётся примерно на одном уровне. Длительность периода зависит от скорости роста обоих видов, а так же от исходных параметров. Для популяции жертвы влияние хищника положительно, т.к. её чрезмерное размножение привело бы к краху её численности. В свою очередь, все механизмы, препятствующие полному истреблению жертвы, способствуют сохранению пищевой базы хищника.
![]()
ГРАФИКИ. Взаимоотношения между хищником и жертвой: А – в гомогенной среде инфузория Dinidium nasatum погибает от голода после поедания жертвы; Б – в гетерогенной среде – жертва частично может укрыться от хищника, и он вымирает; В – естественные колебания численности хищника и жертвы ( по Гаузе).
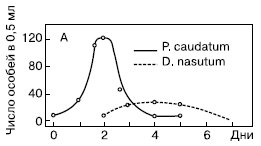
Рис. 3. Взаимодействие между инфузорией дидиниум и инфузорией туфелькой в однородной среде (количество особей в 0,5 см3)
![]()
Рис. 5. Взаимодействие между инфузорией дидиниум и инфузорией туфелькой в неоднородной среде (количество особей в 0,5 см)
![]()
Рис. 6. Взаимодействие между инфузорией дидиниум и инфузорией туфелькой в неоднородной среде при наличии иммиграций (1 инфузория туфелька + 1 дидиниум)
Эксперименты Гаузе.
Жертва – инфузория туфелька Paramecium caudatum.
Хищник – инфузория дидиниум Didinium nasutum.
В качестве пищи для инфузории туфельки Гаузе использовал культуру сенной палочки (Bacillus subtilis).
Первая серия опытов была поставлена Гаузе в так называемой однородной среде. Для этого использовалась разведенная бактериальная культура без осадка. Особенность биологии дидиниум заключается в том, что он не является активным охотником, а просто хватает все, что попадается ему на пути. Инфузория туфелька, ускользая от хищника, использует значительно бóльшую скорость передвижения, а также особые защитные образования – трихоцисты. При этом она никогда не прячется в убежища. Совокупность этих особенностей определила чистоту данной серии опытов и позволило Гаузе выделил процесс взаимодействия хищника и жертвы в чистом виде. Бактериальная культура без осадка разливалась в пробирки объемом 0,5 мл. В нее помещалось 5 особей инфузории туфельки, а через 2 суток вносили 3 особи хищной инфузории дидиниум. Практически сразу же после внесения хищников в культуру, численность жертвы начинала убывать, поскольку дидиниум поедает новую инфузорию туфельку через каждые 3 часа. Дидиниум интенсивно размножался, достигая максимума численности к 3-м – 4-м суткам опыта. На 5-е сутки дидиниум выел всех инфузорий туфелек, а на 6-е – все хищники погибли от голода.
При первой иммиграции в свободную среду дидиниум не нашел жертвы и погиб. После этого начался интенсивный рост инфузории туфельки. К моменту второй иммиграции концентрация инфузории туфельки в среде была уже очень велика, благодаря чему начался рост популяции дидиниум. Третья иммиграция пришлась на момент интенсивного уничтожения жертв хищником, и потому не внесла существенных изменений в общую картину. К четвертой иммиграции дидиниум выел всех инфузорий туфелек, после чего погиб сам. Внесенная в среду инфузория туфелька дала начало новому циклу роста жертв. В дальнейшем, такие изменения численности обоих видов регулярно повторялись.
Проанализировав результаты, полученные в ходе трех серий опытов, Гаузе пришел к заключению, что периодические колебания численности хищника и жертвы не являются свойством процесса взаимодействия между ними, как это предполагали математики. Вероятно, они возникают в результате постоянного вмешательства извне (убежища для жертв, сохранение хищников путем инцистирования, иммиграция и эмиграция, колебания факторов среды и т. д.). Именно поэтому в естественных условиях природа периодических колебаний численности находящихся в непосредственном взаимодействии видов может быть весьма сложной.
Дата добавления: 2015-04-18 ; просмотров: 29 ; Нарушение авторских прав
Паразитизм-ассоциация генетически разнородных организмов, основанная на пищевых связях и взаимообмене, при которой один (паразит) использует другого (хозяина) в качестве среды обитания и источника питания, причем оба партнера находятся в антагонистических отношениях различной степени остроты. Паразит вызывает в организме хозяина иммунобиологические реакции.
Животные организмы, ведущие паразитический образ жизни, принято делить на временных(организмы которые совершают весь цикл развития от яйца до взрослой стадии вне организма хозяина (последнего они используют только для питания) и стационарных(используют хозяина на продолжительное время (или в течение всей его жизни) не только для питания, но и для обитания).
Отношения типа хищник-жертва, паразит-хозяин – это прямые пищевые связи, которые для одного из партнеров имеют отрицательные, а для другого – положительные последствия. Связь хищник– жертва в свою очередь, подразделяются на несколько категорий: 1) истинное хищничество, или хищничество в узком смысле слова; 2) паразитизм; 3) собирательство и 4) пастьба.
Хищниками обычно называют животных, питающихся другими животными, которых они ловят и умерщвляют.
Если размеры жертв намного меньше размеров питающихся ими животных, численность объектов питания высока и сами они легкодоступны – в этом случае деятельность плотоядного вида превращается в поиск и простой сбор добычи и называется собирательство, например, для ряда насекомоядных птиц – куликов-зуйков, ржанок, зябликов, коньков и др.
Паразитизм такая форма связей между видами, при которой организм-потребитель использует живого хозяина не только как источник пищи, но и как место постоянного или временного обитания. По существу, типичный паразитический характер имеют связи насекомых-вредителей с растениями. Паразиты обычно намного мельче своего хозяина.
Во взаимоотношениях хищник-жертва, паразит-хозяин наиболее ярко проявляется эволюционная и экологическая роль пищевых связей организмов. Хищничество, связанное с активным поиском и энергичными способами овладения сопротивляющейся и убегающей добычей, ведет к выработке разнообразных экологических адаптаций как у жертв, так и у их потребителей. При активном способе защиты от врагов естественный отбор способствует развитию у жертв органов чувств, быстроты реакции, скорости бега, инстинктов обманного поведения, что сопровождается совершенствованием нервной системы и ведет к прогрессивной эволюции группы.
При пассивном способе защиты развиваются покровительственная окраска, твердые панцири, шипы, иглы, инстинкты затаивания, использования недоступных хищникам убежищ и т. п. Некоторые из этих способов защиты характерны не только для малоподвижных или сидячих видов, но и для активно спасающихся от врагов животных.
29. Адаптация. Основные пути адаптаций живых организмов к среде
В процессе эволюции у организмов выработались различные приспособления к среде обитания - адаптации. Способность к адаптации — одно из основных свойств живой материи, обеспечивающее возможность ее существования. Адаптации развиваются под действием трех основных факторов: наследственность, изменчивость и естественный (а также искусственный) отбор.
Существует три основных пути приспособления организмов к условиям окружающей среды: активный путь, пассивный путь и избегание неблагоприятных воздействий.
Активный путь — усиление сопротивляемости, развитие регуляторных процессов, позволяющих осуществлять все жизненные функции организма, несмотря на отклонения фактора от оптимума. Например, поддержание постоянной температуры тела у теплокровных животных (птиц и млекопитающих), оптимальной для протекания биохимических процессов в клетках.
Пассивный путь — подчинение жизненных функций организма изменению факторов среды. Например, переход при неблагоприятных условиях среды в состояние анабиоза (скрытой жизни), когда обмен веществ в организме практически полностью останавливается (зимний покой растений, сохранение семян и спор в почве, оцепенение насекомых, спячка позвоночных животных и т.д.).
Избегание неблагоприятных воздействий — выработка организмом таких жизненных циклов и поведения, которые позволяют избежать неблагоприятных воздействий. Например, сезонные миграции животных.
Адаптации можно разделить на три типа: морфологические, физиологические и этологические.
Морфологические адаптации сопровождаются изменением в строении организма (например, видоизменение листа у растений пустынь). Морфологические адаптации у растений и животных приводят к образованию определенных жизненных форм
Физиологические адаптации — изменения в физиологии организмов (например, способность верблюда обеспечивать организм влагой путем окисления запасов жира).
Этологические адаптации - изменения в поведении (например, сезонные миграции млекопитающих и птиц, впадение в спячку в зимний период). Этологические адаптации характерны для животных.
7.3.1. Отношения хищник-жертва, паразит-хозяин
Отношения типа хищник-жертва, паразит-хозяин – это прямые пищевые связи, которые для одного из партнеров имеют отрицательные, а для другого – положительные последствия. По существу, к этому типу экологических взаимодействий можно отнести все варианты пищевых связей. Хотя пасущуюся на лугу корову и дятла, добывающего личинок из-под коры дерева, обычно не называют хищниками, тот тип взаимоотношений, в которые они вступают с организмами, служащими им пищей, имеет много общего с отношениями хищника и его жертв. Поэтому в западной литературе связь хищник– жертва понимается обычно в широком смысле, включая все формы добывания пищи. Они, в свою очередь, подразделяются на несколько категорий: 1) истинное хищничество, или хищничество в узком смысле слова; 2) паразитизм; 3) собирательство и 4) пастьба.
Хищниками обычно называют животных, питающихся другими животными, которых они ловят и умерщвляют. Для хищников характерно специальное охотничье поведение.
Добыча жертвы требует от них значительных затрат энергии на поиск, погоню, захват, преодоление сопротивления жертв.
Если размеры жертв намного меньше размеров питающихся ими животных, численность объектов питания высока и сами они легкодоступны – в этом случае деятельность плотоядного вида превращается в поиск и простой сбор добычи и называется собирательство.
К собирательству можно отнести фильтрационное питание водных животных, седиментацию, или осаждение водной взвеси, сбор пищи илоедами или дождевыми червями. К нему же примыкает так называемое хищничество растений. У многих растений при недостатке азота в питании развиты способы улавливания и фиксации прилетающих к ним насекомых и переваривание белков их тел протеолитическими ферментами (пузырчатки, росянки, непентесы, венерина мухоловка и др.).
По способу овладения пищевыми объектами собирательство приближается к типичной пастьбе фитофагов. Специфика пастьбы заключается в поедании неподвижного корма, находящегося в относительном изобилии, на поиски которого не приходится тратить много усилий. С экологической точки зрения такой способ питания характерен как для стада копытных на лугу, так и для листогрызущих гусениц в кроне дерева или личинок божьих коровок в колониях тлей.
Таким образом, несмотря на экологическую специфичность разных способов питания, они связаны между собой всевозможными переходами, а иногда встречаются вместе у одного и того же вида. Такие же переходы можно встретить в природе между типичным хищничеством и паразитизмом.
Паразитизм такая форма связей между видами, при которой организм-потребитель использует живого хозяина не только как источник пищи, но и как место постоянного или временного обитания (рис. 85). По существу, типичный паразитический характер имеют связи насекомых-вредителей с растениями. Паразиты обычно намного мельче своего хозяина.
![]()
Рис. 85. Паразиты певчих птиц
Во взаимоотношениях хищник-жертва, паразит-хозяин наиболее ярко проявляется эволюционная и экологическая роль пищевых связей организмов. Хищничество, связанное с активным поиском и энергичными способами овладения сопротивляющейся и убегающей добычей, ведет к выработке разнообразных экологических адаптаций как у жертв, так и у их потребителей. При активном способе защиты от врагов естественный отбор способствует развитию у жертв органов чувств, быстроты реакции, скорости бега, инстинктов обманного поведения, что сопровождается совершенствованием нервной системы и ведет к прогрессивной эволюции группы.
При пассивном способе защиты развиваются покровительственная окраска, твердые панцири, шипы, иглы, инстинкты затаивания, использования недоступных хищникам убежищ и т. п. Некоторые из этих способов защиты характерны не только для малоподвижных или сидячих видов, но и для активно спасающихся от врагов животных.
В свою очередь, трудность обнаружения и поимки жертв способствует у хищников отбору на лучшее развитие органов чувств (зоркость, тонкий слух, чутье и т. д.), на более быструю реакцию на добычу, выносливость при преследовании и т. п. Таким образом, экологические связи хищников и жертв направляют ход эволюции сопряженных видов.
Хищники обладают обычно широким спектром питания. Добыча жертв требует много сил и энергии. Специализация ставила бы хищников в сильную зависимость от численности определенного вида жертв. Поэтому большинство видов, ведущих хищный образ жизни, способно переключаться с одной добычи на другую, особенно на ту, которая в данный период более доступна и многочисленна. Правда, у многих хищников есть предпочитаемые виды жертв, которых они добывают чаще, чем других. Эта избирательность может быть обусловлена разными причинами. Во-первых, хищник активно выбирает наиболее полноценную в кормовом отношении пищу. Например, нырковые утки и сиговые рыбы в северных водоемах выбирают среди водных беспозвоночных преимущественно личинок комаров-хирономид (мотыль), и их желудки иногда бывают наполнены мотылем, несмотря на наличие в водоеме и другого корма.
Характер пищи может быть обусловлен также пассивной избирательностью: хищник в первую очередь поедает такие корма, к добыче которых он наиболее приспособлен. Так, многие воробьиные питаются всеми насекомыми, обитающими открыто на поверхности почвы, на траве, листьях и т. п., но не поедают почвенных беспозвоночных, для добычи которых нужны специальные приспособления. Наконец, третьей причиной пищевой избирательности хищников может быть активное переключение на наиболее массовую добычу, появление которой стимулирует охотничье поведение. При высокой численности леммингов даже соколы-сапсаны, основной способ охоты которых – добывание птиц в воздухе, начинают охотиться на леммингов, схватывая их с земли. Способность переключения с одного вида добычи на другой – одно из необходимых экологических приспособлений в жизни хищников.
Паразитизм, в отличие от хищничества, характеризуется более узкой специализацией видов. Поскольку хозяин обеспечивает паразиту не только пищу, но и микроклимат, защиту и т. п., то чем лучше приспособленность паразита к особенностям организма хозяина, тем вероятнее его успех в размножении и оставлении потомства.
Катастрофический вред от паразитов выражен преимущественно лишь в тех связях, которые еще не стабилизированы длительным ходом естественного отбора. Поэтому случайно завезенные вредители поражают сельскохозяйственные растения или животных часто намного сильнее, чем местные.
Среди многообразных форм паразитических отношений есть и такие, при которых гибель хозяина – обязательное следствие пребывания в нем паразита. Этот тип связей особенно распространен у насекомых, откладывающих свои яйца в яйца или личинки других. Такие насекомые получили название паразитоидов. Гибель хозяина обусловлена малым запасом в нем пищи, которой едва хватает на развитие одной или немногих личинок вида-потребителя.
Паразитизм, таким образом, связан всевозможными переходами с другими типами взаимоотношений.
Основная экологическая роль хищничества, паразитизма и других вариантов пищевых связей в сообществах заключается в том, что, последовательно питаясь друг другом, живые организмы создают условия для круговорота веществ, без которого невозможна жизнь. Вторая не менее важная роль этих отношений – взаимная регуляция численности видов.
Данный текст является ознакомительным фрагментом.
Отношения типа хищник — жертва, паразит — хозяин — это прямые пищевые связи, которые для одного из партнеров имеют отрицательные, а для другого — положительные последствия. По существу, к этому типу экологических взаимодействий можно отнести все варианты пищевых связей. Хотя пасущуюся на лугу корову и дятла, добывающего личинок из-под коры дерева, обычно не называют хищниками, тот тип взаимоотношений, в которые они вступают с организмами, служащими им пищей, имеет много общего с отношениями хищника и его жертв. Пищевые взаимосвязи— основные в сообществах. Любой гетеротрофный организм может существовать лишь за счет других гетеротрофных или автотрофных организмов.
Хищниками обычно называют животных, питающихся другими животными, которых они ловят и умерщвляют. Для хищников характерно специально охотничье поведение.
Таким образом, несмотря на экологическую специфичность разных способов питания, они связаны между собой всевозможными переходами, а иногда встречаются вместе у одного и того же вида. Такие же переходы можно встретить в природе между типичным хищничеством и паразитизмом.
Во взаимоотношениях хищник — жертва, паразит — хозяин наиболее ярко проявляется эволюционная и экологическая роль пищевых связей организмов. Хищничество, связанное с активным поиском и энергичными способами овладения сопротивляющейся и убегающей добычей, ведет к выработке разнообразных экологических адаптации как у жертв, так и у их потребителей. При активном способе защиты от врагов естественный отбор способствует развитию у жертв органов чувств, быстроты реакции, скорости бега, инстинктов обманного поведения, что .сопровождается совершенствованием нервной системы и ведет к прогрессивной эволюции группы.
При пассивном способе защиты развиваются покровительственная окраска, твердые панцири, шипы, иглы, инстинкты затаивания, использования недоступных хищникам убежищ и т. п. Некоторые из этих способов защиты характерны не только для малоподвижных или сидячих видов, но отчасти и для активно спасающихся от врагов животных.
В свою очередь, трудность обнаружения и поимки жертв способствует у хищников отбору на лучшее развитие органов чувств (зоркость, тонкий слух, чутье и т. д.), на более быструю реакцию на добычу, выносливость при преследовании и т. п. Таким образом, экологические связи хищников и жертв направляют ход эволюции сопряженных видов.
Хищники обладают обычно широким спектром питания. Добыча жертв требует много сил и энергии. Специализация ставила бы хищников в сильную зависимость от численности определенного вида жертв. Большинство видов, ведущих хищный образ жизни, способно поэтому переключаться с одной добычи на другую, особенно на ту, которая в данный период более доступна и многочисленна. Правда, у многих хищников есть предпочитаемые виды жертв, которых они добывают чаще, чем других. Эта избирательность может быть обусловлена разными причинами. Во-первых, хищник активно выбирает наиболее полноценную в кормовом отношении пищу. Например, нырковые утки и сиговые рыбы в северных водоемах выбирают среди водных беспозвоночных преимущественно личинок комаров-хирономид (мотыль) и их желудки иногда бывают наполнены мотылем, несмотря на наличие в водоеме и другого корма.
Характер пищи может быть обусловлен также пассивной избирательностью: хищник в первую очередь поедает такие корма, к добыче которых он наиболее приспособлен. Так, многие воробьиные питаются всеми насекомыми, обитающими открыто на поверхности почвы, на траве, листьях и т. п., но не поедают почвенных беспозвоночных, для добычи которых нужны специальные приспособления. Наконец, третьей причиной пищевой избирательности хищников может быть активное переключение на наиболее массовую добычу, появление которой стимулирует охотничье поведение. При высокой численности леммингов даже соколы-сапсаны, основной способ охоты которых — добывание птиц в воздухе, начинают охотиться на леммингов, схватывая их с земли. Способ-‘ ность переключения с одного вида добычи на другой — одно из необходимых экологических приспособлений в жизни хищников.
Паразитизм, в отличие от хищничества, характеризуется более узкой специализацией видов. Поскольку хозяин обеспечивает паразиту не только пищу, но и микроклимат, защиту и т. п., то чем лучше приспособленность паразита к особенностям организма хозяина, тем вероятнее его успех в размножении и оставлении потомства.
Катастрофический вред от паразитов выражен преимущественно лишь в тех связях, которые еще не стабилизированы длительным ходом естественного отбора. Поэтому случайно завезенные вредители поражают сельскохозяйственные растения или животных часто намного сильнее, чем местные.
Среди многообразных форм паразитических отношений есть и такие, при которых гибель хозяина — обязательное следствие пребывания в нем паразита. Этот тип связей особенно распространен у насекомых, откладывающих свои яйца в яйца или личинки других. Такие насекомые получили название паразитоидов. Гибель хозяина обусловлена малым запасом в нем пищи, которой едва хватает на развитие одной или немногих личинок вида-потребителя.
Паразитизм, таким образом, связан всевозможными переходами с другими типами взаимоотношений.
Основная экологическая роль хищничества, паразитизма и других вариантов пищевых связей в сообществах заключается в том, что, последовательно питаясь друг другом, живые организмы создают условия для круговорота веществ, без которого невозможна жизнь. Вторая не менее важная роль этих отношений — взаимная регуляция численности видов.
В лабораторных условиях при совместном содержании хищника и его жертвы отмечаются периодические изменения численности обоих видов, имеющие довольно закономерный характер. В опытах, проведенных нашим отечественным биологом Г. Ф. Гаузе в 30-х годах с пробирочными культурами простейших, было обнаружено следующее. Усиленное размножение хищной инфузории Didinium nasutum закономерно следовало всякий раз за размножением ее жертвы — туфельки Paramecium cauda-tum. Когда хищник достигал высокой численности, он полностью уничтожал парамеций, после чего наступала гибель самих Didinium, лишенных пищевой базы. Однако когда в пробирки помещали некоторое количество песчинок, под которым часть туфелек могла найти недоступные для хищников убежища, после гибели Didinium наступало новое массовое размножение парамеций. В другой серии опытов, когда культивировались разные виды парамеций, а- в качестве их пищи дрожжевне клетки рода Saceharomyces, было получено несколько закономерных циклов колебаний численности дрожжей и туфелек. Механизмы подобных колебаний можно представить следующим образом.
![]()
Увеличение численности жертв происходит до тех пор, пока темп выедания их нарастающим количеством хищников не превысит скорости размножения. После этого дальнейшая деятельность хищников только подрывает их кормовую базу, что ведет к снижению их собственной численности. Это позволяет немногим оставшимся в живых жертвам вновь размножиться и, следовательно, вновь улучшить кормовые условия для хищников. Если последние еще присутствуют в системе, новое увеличение количества жертв способствует новому подъему их численности и цикл повторяется снова.
В природных условиях такие закономерные циклические изменения численности двух видов, связанных пищевыми отношениями, выявить труднее так как на их размножение и гибель влияют взаимодействия со множеством других видов, а также абиотические изменения среды. Однако в ряде случаев и в природной обстановке удается заметить регулярные периодические изменения обилия хищников и их жертв. При этом прослеживаются следующие основные закономерности.
Общее число жертв, которых может уничтожить один хищник, сначала растет почти пропорционально росту численности потребляемого вида. Это так называемая функциональная реакция хищников на жертвы. Однако она имеет предел, обусловленный физическими возможностями потребителя. После полного насыщения хищники обычно слабо реагируют на жертву. Дальнейшее повышение истребляемости жертв достигается увеличением численности хищников за счет- успешного размножения на хорошей кормовой базе. Это явление получило название численной или количественной реакции хищников на рост численности жертв. Количественная реакция хищников всегда запаздывает по сравнению с ходом размножения жертвы, так как является ответом на изменение кормовой базы. Эти сдвиги усиливают цикличность колебаний.
![]()
Периоды возникающих колебаний численности относительно постоянны, тогда как амплитуда их может изменяться в широких пределах в связи с влиянием других факторов среды.
Колебания продолжаются до тех пор, пока в результате различных (чаще случайных) причин численность одного из партнеров не упадет до нуля.
Особенно наглядно такие колебания обнаруживаются в обедненных сообществах, где межвидовые связи не столь разнообразны: в тундрах и полярных пустынях, лесах с господством одной породы деревьев, культурных садах и т. п. Циклические изменения численности выявлены, например, у леммингов и их основных потребителей — песцов, массово размножающихся хвое- и ли-стогрызущих насекомых и их паразитов, вредителей яблонь — паутинных клещиков и питающихся ими хищных клещей и у многих других сопряженных пищевыми связями видов.
Таким образом, истребление жертв и поражаемость хозяев Зависят от численности врагов, но и обилие последних также определяется численностью объектов их питания.
Читайте также:
 Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу.
Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу.
Copyright © Иммунитет и инфекции