
Мовні центри кори головного мозку
Можливість існування характерних ознак поведінки, що здійснюється за участю сигнальних систем, знайшла згодом підтвердження у наявності функціональної асиметрії двох великих півкуль головного мозку, які діють як одне ціле. Виключення ОДНІЄЇ з них дозволяє виявити представництво функцій, характерних для особливостей поведінки художнього чи розумового типів ВНД людини.
Ліву півкулю називають домінуючою, бо її функції пов'язані з мовою людини як у правші, так і в шульги; пошкодження цієї півкулі призводить до втрати мови – афазії.
Існує два центри мови – Верніке і Брока, різних за своїм функціональним призначенням. Пошкодження сенсорного центру Верніке (розміщений в корі задньої частини верхньої скроневої звивини) призводить до сенсорної афазії – людина не розуміє сказані чи написані слова, але може розмовляти.
Пошкодження рухового центру Брока (знаходиться у корі заднього відділу лобової звивини, поблизу зони, що іннервує лицеву мускулатуру) призводить до моторної афазії – людина не може писати і говорити, але розуміє слова. Центри Брока та Верніке пов'язані між собою дугоподібними зв'язками. Вважають, що центр Верніке є первинним центром мови. Видалення його призводить до суттєвіших порушень мови, ніж видалення центру Брока (рис. 8.13).
Права півкуля на відміну від лівої домінує у проявах емоцій (міміка, інтонація, жестикуляція – "мова тіла"), розв'язанні просторових задач, музикальному сприйнятті, інтуїції. Відомий французький композитор Моріс Равель у 57 років отримав травму лівої півкулі при автомобільній катастрофі. Він, як і до того, міг слухати музику, отримувати задоволення, але не зміг займатися композицією – записати те, що звучало в його голові. У нього виникла афазія Брока.
Спеціалізація півкуль також пов'язана з переважаннями функції правої чи лівої руки, що є генетично обумовленим. У 96 % правшів (91 % людства) домінує ліва півкуля, у решти 4 % – права.
Функціональна асиметрія (латералізація) півкуль великого мозку
Мозок діє як єдине ціле, хоча має дві півкулі. Інформація передається між двома півкулями кори головного мозку через мозолисте тіло і мозок функціонує як єдина структура, сумісно здійснюючи всі функції. Винятком є передня частина скроневої частки. Ця скронева структура, в тому числі мигдалина, взаємодіють через нервові волокна передньої комісури. У дитячому віці мозок має значну пластичність і може компенсувати пошкодження структур. Пошкодження мозолистого тіла припиняє передачу інформації від центру Верніке домінуючої півкулі до моторної кори протилежної півкулі мозку, що призводить до втрати контролю правої рухової кори над свідомими руховими функціями лівої кисті й передпліччя, навіть тоді, коли в нормальних умовах вони є підсвідомими (рис. 8.14).
Переріз мозолистого тіла спричиняє передачу соматичної й зорової інформації від правої півкулі до лівої

РИС. 8.13. Карта специфічних функціональних ділянок, на яких виділені ділянки Верніке і Брока, відповідальні за розуміння та відтворення мови; у 95% людей вони локалізовані в лівій півкулі

РИС. 8.14. Функціональна асиметрія півкуль головного мозку людини: А - функції правої і лівої півкуль; Б - правильне зображення; В. Г - відтворення хреста і паралелепіпеда людиною з "розщепленим" мозком лівою (В) і правою (Г) рукою
півкулі поля Верніке. Таким чином, існує дві свідомих частки мозку. Наприклад, лише завдяки лівій півкулі мозку людина розуміє як написані, так і сказані слова. Права півкуля може розуміти лише написані слова та не розуміє сказаних.
У більшості людей права півкуля переробляє інформацію в цілому, а в лівій півкулі здійснюється її послідовна переробка з використанням слів – мови. Мова – це унікальний вид переробки інформації, властивий лише людині. Завдяки властивостям кори головного мозку людство досягло технічного прогресу й розвитку культури.
Кора головного мозку: зони- сенсорна,асоціативна,рухова. Кора головного мозку: зона сенсорна
Сенсорні зони – це функціональні зони кори головного мозку, які через висхідні нервові шляхи отримують сенсорну інформацію від більшості рецепторів тіла. Вони займають окремі ділянки кори, пов’язані з певними видами відчуттів. Розміри цих зон корелюють з числом рецепторів у відповідній сенсорній системі.
– Первинні сенсорні зони і первинні моторні зони (проекційні зони);
– Вторинні сенсорні зони і вторинні моторні зони (асоціативні одномодальних зони);
– Третинні зони (асоціативні разномодальних зони);
Первинні сенсорні і моторні зони займають менше 10% поверхні кори головного мозку і забезпечують найбільш прості сенсорні і рухові функції.
Кора головного мозку: зони асоціативні (функціональні)
Асоціативні зони – це функціональні зони кори головного мозку. Вони пов’язують знову що надходить сенсорну інформацію з полученой раніше і зберігається в блоках пам’яті, а також порівнюють між собою інформацію, що отримується від різних рецепторів. Сенсорні сигнали інтерпретуються, осмислюються і при необхідності використовуються для визначення найбільш підходящих відповідних реакцій, які вибираються в асоціативної зоні і передаються в пов’язану з нею рухову зону. Таким чином, асоціативні зони беруть участь у процесах запам’ятовування, навчання і мислення, і результати їх діяльності складають те, що зазвичай називають інтелектом.
Окремі великі асоціативні області розташовані в корі поруч з відповідними сенсорними зонами. Наприклад, зорова асоціативна зона розташована в потиличній зоні безпосередньо попереду сенсорної зорової зони і здійснює описані вище асоціативні функції, пов’язані із зоровими відчуттями. Деякі асоціативні зони виконують лише обмежену спеціалізовану функцію і пов’язані з іншим асоціативними центрами, здатними піддавати інформацію подальшій обробці. Наприклад, звукова асоціативна зона аналізує звуки, розділяючи їх на категорії, а потім передає сигнали в більш спеціалізовані зони, такі як мовна асоціативна зона, де сприймається сенс почутих слів.
Ці зони відносяться до асоціативної корі і беруть участь в організації когнітивних функцій і складних форм поведінки.
– Вторинні сенсорні зони і вторинні моторні зони (асоціативні одномодальних зони);
– Третинні зони (асоціативні разномодальних зони);
– Паралімбіческіе зони і
Кора головного мозку: зона рухова
Рухові зони – це функціональні зони кори головного мозку, що посилають рухові імпульси до довільних м’язам по спадним шляхах, які починаються в білій речовині великих півкуль.
Багато рухові імпульси йдуть прямо в спинний мозок через два великих пірамідних тракту (кортікоспінальних тракту), що проходять в стовбурі мозку. Решта рухові імпульси передаються по екстрапірамідних шляхів, тут же йдуть рухові імпульси від базальних гангліїв і мозочка. У довгастому мозку всі шляхи перехрещуються, так що імпульси, які йдуть від кори лівої півкулі, іннервують праву половину тіла і навпаки.
Тіла нейронів, що у освіті пірамідних трактів, лежать в рухових зонах кори, а їх аксони утворюють синапси безпосередньо з мотонейронами спинного мозку в тому його сегменті, де ці нейрони виходять на периферію. У головному мозку немає ніяких проміжних синапсів, тому імпульси і наступні відповіді на них по шляху не затримуються і не видозмінюються.
Головним екстрапірамідних трактом є ретікулоспінальний тракт, перемикаючий імпульси від ретикулярної формації, яка лежить в стовбурі мозку між таламуса і довгастим мозком. З різних відділів головного мозку, що контролюють рухову активність, імпульси надходять у певні ділянки ретикулярної формації, де вони модифікуються під впливом імпульсів, що йдуть від кори, і стають або збудливими, або гальмівними. Наприклад, імпульси від мозочка і премоторной зони кори, керуючої координованими рухами, надходять в ту область ретикулярної формації, яка знаходиться в довгастому мозку і посилає імпульси, що стимулюють гальмівні мотонейрони. Останні придушують активацію певних м’язів, що дає можливість здійснювати складні координовані рухи тіла. Інші комбінації рухових імпульсів, навпаки, стимулюють збуджуючі нейрони, і загальний вплив ретикулярної формації на рухову активність виявляється збудливим.
Більшість волокон сенсорних нейронів на своєму шляху через таламус до кори віддає колатералі (бічні гілки) у ретикулярну формацію, беручи участь в утворенні ретикулярної активує системи, яка тонізує кору і бере участь в пробудженні організму від сну. Недостатня активність цієї системи або її руйнування приводить відповідно до глибокого сну або коми. Як вважають, багато речовини, що викликають загальний наркоз, надають свою дію, тимчасово блокуючи синаптичну передачу в цій системі. Передбачається також, що ретикулярна активує система відповідальна за виникнення і підтримання спонукань до дії і концентрації уваги.
Роль кори головного мозку в системній організації діяльності організму
1. Структурно-функціональні особливості кори
2. Роль кортикальних колонок
3. Проекційні та асоціативні поля
4. Методи вивчення кори великих півкуль
Кора великих півкуль з’являється вперше у риб у вигляді нюхових клітин, але видалення їх суттєво не впливає на поведінку риб. В амфібій вже є нюхова кора. Півкулі з’являються лише у птахів, у них при видаленні півкуль помітно змінюється поведінка: самі не можуть злетіти, самі не можуть їсти, в’ють гнізда, але не реагують на крик пташенят, у них зникає материнський інстинкт. Повного розвитку кора досягає у ссавців. У них видалення кори викликає втрату материнського інстинкту (собаки можуть поїдати своїх щенят), тварини не обходять перепони, натикаються на предмети, не йдуть на клич господаря, не орієнтуються в просторі, у них зникає захисний інстинкт. Найбільшу площу займає кора у приматів – 2200 см 2 . В них найкраще розвинені борозни та звивини. Функції кори повністю забезпечують пристосування до життя та вищу психічну діяльність. Видалення великих півкуль призводить до втрати здатності до самостійного життя.
У людини кора великих півкуль забезпечує такі функції:
- Взаємодія організму з навколишнім середовищем.
- Регуляція діяльності внутрішніх органів.
- Регуляція обміну речовин та енергії.
- Вища нервова діяльність –мова, пам’ять, мислення, свідомість.
Кора головного мозку є вищим відділом ЦНС. Це сіра речовина товщиною 3-5 мм, вкриває півкулі головного мозку. Вона займає площу 22 м 2 , утворюючи багаточисельні складки. В складі кори до 10 9 -10 10 нейронів, які утворюють 6 шарів:
1. Молекулярний шар – має мало клітин, їх волокна утворюють поверхневе густе тангенціальне сплетіння з дендритами другого шару.
2. Зовнішній зернистий шар – пірамідні клітини середньої величини, волокна яких розташовані радіально.
3. Внутрішній зернистий шар – зірчасті клітини, волокна яких розташовані горизонтально.
4. Внутрішній пірамідний (гангліозний) шар – це гігантські пірамідні клітини Беца, які мають довгі дендрити, що тягнуться до молекулярного шару.
5. Поліморфний шар – це шар веретеноподібних клітин.
Зв’язок кори великих півкуль з підкорковими структурами здійснюється за допомогою аферентних і еферентних волокон. Аферентні волокна називаються кортикопетальними , вони несуть інформацію в кору. Основними з них є таламокортикальн волокна . Це прямі аферентні шляхи, які розгалужуються у внутрішньому зернистому шарі і не дають колатералей. Невелика частина волокон йде в молекулярний шар, утворюючи колатеральні еферентні волокна, які називаються кортикофугальними , вони несуть інформацію від кори до підкоркових структур. Ці волокна діляться на 3 групи:
1. Проекційні – прямі еферентні волокна, що утворюють провідні шляхи (кортикоспінальні, кортикоталамічні)
2. Асоціативні - волокна, що утворюють безліч колатералей та йдуть в різні підкоркові зони однойменної півкулі.
3. Комісуральні - волокна, що йдуть від кори в складі мозолистого тіла і з’єднують зони кори однієї півкулі з підкорковими зонами другої.
1, 2 шари кори великих півкуль забезпечують аналіз та синтез отриманої інформації, мають багато асоціативних волокон.
3, 4 шари кори великих півкуль одержують інформацію від усіх органів та частин тіла за рахунок кортикопетальних волокон.
5, 6 шари кори великих півкуль – це рухові нейрони, звідси починаються рухові шляхи, що включають кортикофугальні волокна.
В шарах клітини розміщуються перпендикулярно до поверхні кори, утворюючи ланцюги. Елементарні нервові ланцюги відповідають за переробку певної інформації. Такий функціональний принцип названо – кортикальні колонки. Це елементарна функціональна одиниця, в якій здійснюється локальна переробка інформації від рецепторів однієї модальності. Кожна колонка має діаметр 500-1000 мкм, в складі яких розміщується 5-6 нейронів. Пірамідні клітини орієнтовані вертикально, їх аксони утворюють зворотні колатералі, які забезпечують як процеси полегшення в межах мікромодуля, так і гальмування між мікромодулями. Аксони зірчастих клітин ідуть через інтернейрони горизонтально, тому вони, головним чином, забезпечують гальмівні процеси. Веретеноподібні клітини мають довгі аксони, які орієнтовані як горизонтально, так і вертикально. Вони формують кортико-таламічні шляхи.
Мікромодулі об’єднуються в макромодулі завдяки горизонтальним розгалуженням терміналей. В колонці можуть бути прості та складні нейрони. Поряд з цим, в корі існує система, яка зчитує елементарні процеси в колонках та об’єднує всю інформацію. Формування таких систем зумовлено внутрішньо-кортикальними зв’язками між окремими макромодулями. Збудження одного мікромодуля викликає гальмування сусідніх. Активація мікромодулів відбувається за рахунок горизонтальних волокон таламокортикальних шляхів.
За функціями поля кори великих півкуль неоднозначні. Вивчення ролі окремих зон кори методом подразнення стало основою для вчення про локалізацію функцій в корі великих півкуль. Пізніше І.П. Павлов методом видалення в поєднанні з методом умовних рефлексів, підтвердив локалізацію функцій в корі. Разом з тим, Лешлі та ін. вважали, що різні зони кори великих півкуль є еквіпотенціальними, тобто однаковими за функціями – так виникла теорія еквіпотенціальності. Така думка була помилковою, бо тільки у нижчих тварин кора не має спеціалізованих зон. Так Лешлі, зруйнувавши кору щурів, знайшов, що порушення навиків руху щурів по лабіринту не залежить від місця руйнування. Досліди на тваринах з використанням мікроелектродного методу показали, що зони кори неоднозначні за функціями. Проекційні поля, створені мономодальними нейронами, одержують інформацію від рецепторів через переключаючі ядра таламуса. Вони оцінюють вид подразнення. На їх долю припадає 14% кори. Це специфічні проекційні поля. Асоціативні поля (86%) знаходяться навколо проекційних полів. Це полімодальні нейрони, які одержують інформацію від асоціативних ядер таламуса. Вони оцінюють джерело подразнення, його властивості та відстань до нього. Серед асоціативних полів є вищі поля, які забезпечують психічну діяльність людини, а також є поля, що формують пам’ять, індивідуальний досвід, прогнозування, передбачення. Це орбіто-фронтальні поля та тім’яна кора. Ці поля співпадають з функціональними зонами, де знаходяться нервові центри.
Представництво всіх видів чутливості в задній центральній звивині.
Задня центральна звивина – поля по Бродману – 1, 2, 3 – корковий центр всіх видів шкірної та пропріоцептивної чутливості. Вона має такі особливості:
1. Проекція протилежної сторони тіла розміщена вниз головою.
2. Має місце соматотопічна організація для кожної частини тіла.
3. Поля шкірної чутливості неоднозначні за площею, так, проекційні поля для обличчя, губ, кистей рук займають найбільшу площу.
При подразненні цієї зони виникає відчуття оніміння, тиску, повзання мурашок. При руйнуванні її зникає чутливість на протилежній стороні – анестезія.
Проекція рухових центрів в передній центральній звивині.
Передня центральна звивина – поля по Бродману – 4, 6 – моторна зона. Вона забезпечує згинання, розгинання, пронацію, супінацію, ротацію. Вона також має соматотопічну організацію, але тут найбільшу площу займають мімічні м’язи обличчя, язик, кисті рук. При подразнення цієї зони виникає скорочення м’язів протилежної сторони тіла. При пошкодженні – розвиваються парези та паралічі. Парез – це обмеження довільних рухів. Параліч – це відсутність рухів.
Сенсорні системи мають подвійну організацію:
а) SS1 – це сомато-сенсорна зона – І, знаходиться в задній центральній звивині, виконує дискримінативний аналіз, тобто оцінює форму і характер поверхні предмету, приймає участь в складних рухах;
б) SS2 – це сомато-сенсорна зона – ІІ, має відношення до формування регуляції довільних рухів, орієнтації голови у напрямку звуків. Обидві зони мають сенсорні та моторні входи.
в) SМ1 – це сомато-моторна зона – І, знаходиться в передній центральній звивині;
г) SМ2 – це сомато-моторна зона – ІІ, знаходиться там же на медіальній поверхні кори. Обидві зони забезпечують відчуття просторової протяжності, стереогноз, відчуття ваги.
Потилична кора – це корковий центр зору. Він оцінює вигляд, форму, розміри та колір предмету. При пошкодження виникає геміанопсія (випадає частина поля зору), або виникає зорова агнозія (людина не впізнає знайомі предмети).
Верхня скронева звивина – корковий центр слуху (звивина Гешля – поля 41, 42). Оцінює звукові подразнення. При пошкодженні людина не може повторити сказане слово, хоч і чує, людина не розуміє звернену до неї мову, це слухова агнозія.
Задня частина нижньої лобної звивини (центр Брока) – це моторний центр мови. У правців він розміщується зліва. При пошкодження людина втрачає здатність до виразного мовлення, узгодження слів у реченні – моторна афазія.
Задня частина верхньої лобної звивини (центр Верніке) – це сенсорний центр мови, відповідає за сприйняття мови. При пошкодженні людина не розуміє звернену до неї мову – сенсорна афазія.
Задній відділ середньої лобної закрутки – руховий центр письма. При пошкодженні людина втрачає здатність писати – аграфія.
Кутова закрутка нижньої тім’яної долі (поле 30) – центр читання. При пошкодженні людина втрачає здатність читати – алексія.
Морський коник та його гачок – центр нюху. При пошкодженні людина втрачає здатність розрізняти запахи.
Верхні тім’яні долі – центр стереогнозу – оцінка положення тіла в просторі з закритими очима, глибока чутливість.
Надкрайова звивина (поле 40 лівої півкулі) – забезпечує здатність здійснювати привичні рухи чи автоматичні рухи, що сформувались в процесі життя. При пошкодженні розвивається моторна апраксія – людина знає, як виконати рух. але не може зробити його, не може намалювати предмет (порушується послідовність виконання складових руху).
Методи вивчення функцій кори великих півкуль:
1. Анатомо-клінічний метод – спостерігають за поведінкою людини при житті, вивчають структурні зміни після смерті.
2. Порівняльно-фізіологічний – оцінюють роль кори по поведінці у тварин різного рівня еволюційного розвитку.
3. Гістологічний – вивчають особливості будови кори на різних етапах розвитку організму.
4. Метод подразнення – вивчають функції кори шляхом подразнення різних її відділів.
5. Метод часткового та повного видалення кори – вивчають зміни поведінки тварин після видалення частин кори. (Роландо, 1823 рік).
6. Метод умовних рефлексів – І.П. Павлов вивчав поведінкові реакції у собак при сполученні різних подразників.
Генерацію електричних коливань в корі великого мозку живих організмів спостерігали і вперше описали Кеннон Р. (1875) і Данилевський В.Я. (1876). Реєстрація електричних коливань потенціалу від поверхні голови була проведена вперше в 1925 р. Правдич-Неминським В.В. у дослідах на тваринах і німецьким психіатром Н. Berger (1929) у людей.
У людей електрична активність кори великого мозку відводиться звичайно від шкіри на черепі. Метод реєстрації постійної електричної активності кори великого мозку отримав назву електроенцефалографія.
Для одержанная електроенцефалограми (ЕЕГ) можна застосувати або біполярний запис від двох активних електродів, розташованих на шкірі голови, або монополярний запис, при якому реєструючий (активний) електрод фіксують на шкірі голови, а індиферентний – на деякій відстані від нього (наприклад, на мочці вуха).
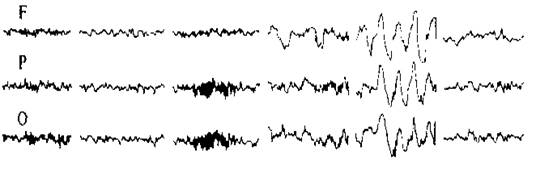
Запис ЕЕГ проводять за допомогою електроенцефалографу. При аналізі ЕЕГ враховують насамперед частоту, амплітуду, форму, тривалість та характер розподілу хвиль. Аналіз можна проводити візуально і за допомогою спеціальних електронних аналізаторів (аналогових і цифрових приладів). На сьогодні детально описано характеристики ЕЕГ для різних станів кори, точно виміряне і проаналізовано коливання потенціалів, які її складають. У несплячої людини в розслабленому стані при закритих очах за відсутності будь-яких зовнішніх подразників переважають хвилі частотою 8-13 Гц, амплітудою близько 50-100 мкВ, виражені вони більшою мірою в потиличних частках кори. Такі хвилі називають альфа-хвилями (або основним ритмом). Це так звана синхронізована ЕЕГ.
Коли людина розплющує очі, альфа-хвилі зникають (так звана блокада альфа-ритму або реакція десинхронізації) і замість них виникають бета-хвилі, які характеризуються більшою частотою (14-30 Гц) і меншою (до 30 мкВ) амплітудою. Вони відображають стан активності мозку, тобто збудження.
Під час перехідного стану від спокою до сну з’являється повільно-хвильовий ритм амплітудою 100-150 мкВ, частотою 4-7 Гц. Це тета-ритм. Під час глибокого сну на ЕЕГ реєструється дельта-ритм амплітудою 250-350 мкВ, частотою 0,5-4 Гц.
F - лобна кора, Р – тім’яна кора, О – потилична кора.
бета-ритм альфа-ритм тета-ритм дельта-ритм
ЕЕГ відображає головним чином постсинаптичні потенціали нейронів кори великого мозку. Про це свідчать результати одночасної реєстрації ЕЕГ за допомогою поза- і внутрішньоклітинного запису активності кортикальних нейронів. Припускають, що позитивні коливання потенціалів на поверхні кори великого мозку пов’язані з ЗПСП, який виникає в нейронах глибинних шарів кори або ГПСП у поверхневих її шарах.
Негативні коливання потенціалів ЕЕГ зумовлені зворотніми процесами в цих шарах. Ритмічна активність кори великого мозку, зокрема альфа-ритм, зумовлена головним чином впливами підкіркових структур, особливо таламуса, в якому виявлено особливі нейрони – “водії ритму” (пейсмекери), котрі через відповідні збудливі і гальмівні зв’язки здатні генерувати і підтримувати ритмічну активність кори великого мозку.
Запис ЕЕГ– діагностична процедура, яка використовується в неврологічній практиці. Так, при дифузних органічних пошкодженнях головного мозку, черепно-мозкових травмах спостерігаються сповільнені і нерегулярні хвилі. При пухлинах мозку часто виникають місцеві зміни ЕЕГ (у ділянці пухлин). У хворих на епілепсію на ЕЕГ спостерігаються пароксизмальні потенціали, судомні розряди, гострокінцеві хвилі та інші зміни. Запис ЕЕГ широко використовується в хірургічній практиці для контролю глибини наркозу: під час глибокої стадії наркотичного сну на ЕЕГ переважають дельта-хвилі.
При констатації смерті у сумнівних випадках, особливо при реанімації хворого, в клініці часто орієнтуються на зникнення коливань на ЕЕГ. (“плоска ЕЕГ”). У клінічній практиці також використовують метод реєстрації викликаних потенціалів для одержання об’єктивних даних про характер і динаміку деяких порушень сенсорних функцій.
При подразненні рецепторів шкіри, м’язів у первинних сомато-сенсорних ділянках переключаючих ядер таламуса кори великих півкуль реєструють коротко-латентні позитивно-негативні коливання електричного потенціалу, які носять назву первинні викликані потенціали. Вони постійні за формою, мають ЛП в межах 1-15 мсек, стійкі до наркозу та негативних чинників, здатні ритмічно відтворюватись. Зі значно більшим латентним періодом і більш складної форми реєструються потенціали, викликані подразненням неспецифічних висхідних шляхів, вони називаються вторинними потенціалами. Вторинні потенціали реєструються тільки в асоціативній корі, мають ЛП більше 15 мсек, зникають при поглибленні наркозу, нестабільні за формою, ритмічне відтворення їх одержати нелегко. Реєстрація викликаних потенціалів широко використовується для вивчення локалізації функцій в корі великих півкуль та встановлення зв’язків між різними структурами ЦНС.
На підставі численних досліджень з певною точністю встановлено функціональне значення різних областей кори півкуль великого мозку.
Ділянки кори півкуль, що мають характерну цитоархітектоніку, і нервові зв'язки, що беруть участь у виконанні певних функцій, є нервовими центрами. Поразка таких ділянок кори виявляється у втраті притаманних їм функцій. Нервові центри кори півкуль великого мозку можуть бути розділені на проекційні і асоціативні.
Проекційні центри - це ділянки кори півкуль великого мозку, що представляють собою корковую частина аналізатора, що мають безпосередній морфофункціональну зв'язок через аферентні або еферентні провідні шляхи з нейронами підкоркових центрів. Вони здійснюють первинну обробку надходить свідомої афферентной інформації та реалізацію усвідомленої еферентної інформації (довільні рухові акти).
Асоціативні центри - це ділянки кори півкуль великого мозку, що не мають безпосереднього зв'язку з підкірковими утвореннями, а пов'язані тимчасової двостороннім зв'язком з проекційними центрами. Асоціативні центри відіграють першорядну роль у здійсненні вищої нервової діяльності (глибока обробка свідомої афферентной інформації, розумова діяльність, пам'ять і т.д.).
В даний час досить точно з'ясована динамічна локалізація деяких функцій кори півкуль великого мозку.
Ділянки кори півкуль великого мозку, які не є проекційними або асоціативними центрами, беруть участь у виконанні межаналізаторних інтегративної діяльності головного мозку.
Проекційні нервові центри кори півкуль великого мозку розвиваються як у людини, так і у вищих хребетних тварин. Вони починають функціонувати відразу ж після народження. Формування цих центрів завершується набагато раніше, ніж асоціативних. У практичному відношенні важливими є наступні проекційні центри.
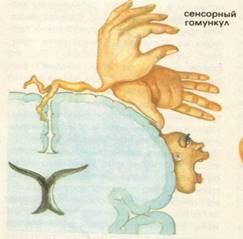
1. Проекційний центр загальної чутливості (тактильної, больової, температурної і свідомої пропріоцептивної) також називають шкірним аналізатором загальної чутливості. Він локалізується в корі постцентральной звивини (поля 1, 2, 3). У ньому закінчуються волокна, що йдуть у складі таламо-коркового шляху. Кожна ділянка протилежної половини тіла має виразну проекцію в кірковій кінці шкірного аналізатора (соматотопическую проекція). У верхньому відділі постцентральна звивини проектуються нижня кінцівку і тулуб, в середньому - верхня кінцівка і в нижньому - голова (сенсорний гомункулюс Пенфілда). Розміри проекційних зон соматосенсорной кори прямо пропорційні кількості рецепторів, що у шкірних покривах. Цим пояснюється наявність найбільш великих соматосенсорних зон, відповідних обличчю і кисті (рис. 3.25). Поразка постцентральна звивини викликає втрату тактильної, больової, температурної чутливості і м'язово-суглобового відчуття на протилежній половині тіла.

Мал. 3.25. Соматотопическую проекція зон чутливості на постцентральну звивину (сенсорний гомункулюс Пенфілда):
- 1 - статеві органи; 2 - стопа; 3 - стегно; 4 - тулуб; 5 - кисть; 6 - вказівний і великий пальці кисті; 7 - особа; 8 - зуби; 9 - мова; 10 - глотка і внутрішні органи
- 2. Проекційний центр рухових функцій (кінестетичний центр), або руховий аналізатор, розташовується в руховій області кори, що включає перед- Центральая звивину і навколоцентральні часточку (поля 4, 6). У 3-4-му шарах кори рухового аналізатора закінчуються волокна, що йдуть у складі таламо-коркового шляху.
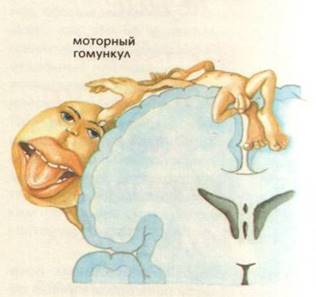
Тут здійснюється аналіз пропріоцептивних (кінестетичних) подразнень. У п'ятому шарі кори розташовується ядро рухового аналізатора, від нейроцитів якого беруть початок корково-спинномозкової і корково-ядерний шляху. У предцентральна звивині також є чітка соматотопическую локалізація рухових функцій. М'язи, виконують складні і тонко диференційовані рухи, мають велику проекційну зону в корі предцентральной звивини. Найбільшу площу займає проекція м'язів язика, обличчя і кисті, найменшу - проекція м'язів тулуба та нижніх кінцівок. Соматотопическую проекція на предцентральна звивину носить назву "моторний гомункулюс Пенфілда". Тіло людини проектується на звивині "догори ногами", причому проекція здійснюється на кору протилежної півкулі (рис. 3.26).
Аферентні волокна, що закінчуються в чутливих шарах кори кінестетичного центру, спочатку проходять у складі шляхів Голля, Бурдаха і ядерно-таламического тракту, проводять імпульси свідомої пропріоцептивної чутливості. Поразка предцентральной звивини призводить до порушення сприйняття роздратувань від скелетних м'язів, зв'язок, суглобів і окістя. Корково-спинномозковою та корково-ядерний шляху проводять імпульси, що забезпечують свідомі руху, і надають гальмівне вплив на сегментарний апарат стовбура головного і спинного мозку. Корковий центр рухового аналізатора через систему асоціативних волокон має численні зв'язки з різними корковими сенсорними центрами (центром загальної чутливості, центром зору, слуху, вестибулярних функцій і т.д.). Зазначені зв'язку необхідні для виконання інтегративних функцій при виконанні довільних рухів.
3. Проекційний центр схеми тіла розташовується в області внутритеменной борозни (поле 40s). У ньому представлені соматотопическую проекції всіх частин тіла. У центр схеми тіла надходять імпульси переважно свідомої пропріоцептивної чутливості. Основне функціональне призначення даного проекційного центру - визначення положення тіла і окремих його частин у просторі і оцінка тонусу мускулатури. При ураженні верхньої тім'яної часточки спостерігається порушення таких функцій, як впізнавання частин власного тіла, відчуття зайвих кінцівок, порушення визначення положення окремих частин тіла в просторі.

Мал. 3.26. Соматотопическую проекція рухових зон на предцентральна звивину (моторний гомункулюс Пенфілда):
- 1 - стопа; 2 - гомілка; 3 - коліно; 4 - стегно; 5 - тулуб; 6 - кисть; 7 - великий палець кисті; 8 - шия; 9 - особа; 10 - губи; 11 - мова; 12 - гортань
- 4. Проекційний центр слуху, або ядро слухового аналізатора, розташовується в середній третині верхньої скроневої звивини (поле 22). У цьому центрі закінчуються волокна слухового шляху, що відбуваються від нейронів медіального колінчастого тіла (підкірковий центр слуху) своєї і, переважно, протилежної сторін. У кінцевому рахунку волокна слухового шляху проходять у складі слуховий лучистости.
При ураженні проекційного центру слуху з одного боку відзначається зниження слуху на обидва вуха, причому з протилежного боку від осередку ураження слух знижується більшою мірою. Повна глухота спостерігається тільки при двосторонній поразці проекційних центрів слуху.
5. Проекційний центр зору, або ядро зорового аналізатора, локалізується на медіальній поверхні потиличної частки, по краях шпорної борозни (поле 17). У ньому закінчуються волокна зорового шляху зі своєю і протилежною сторін, що відбуваються від нейронів латерального колінчатого тіла (підкірковий центр зору). На шпорная борозну є певна соматотопическую проекція різних ділянок сітківки.
Однобічне ураження проекційного центру зору супроводжується часткової сліпотою на обидва ока, але в різних ділянках сітківки. Повна сліпота настає тільки при двосторонній поразці.
- 6. Проекційний центр нюху, або ядро нюхового аналізатора, розташовується на медіальній поверхні скроневої частки в корі парагиппокампальной звивини і в гачку. Тут закінчуються волокна нюхового шляху зі своєю і протилежною сторін, що відбуваються від нейронів нюхового трикутника. При односторонньому ураженні проекційного центру нюху відзначаються зниження нюху і нюхові галюцинації.
- 7. Проекційний центр смаку, або ядро смакового аналізатора, розташовується там же, де і проекційний центр нюху, тобто в лімбічної області мозку (гачок і парагіппокампальная звивина). У проекційному центрі смаку закінчуються волокна смакового шляху своєї і протилежної сторін, що відбуваються від нейронів базальних ядер таламуса. При ураженні лімбічної області спостерігаються розлади смаку, нюху, нерідко з'являються відповідні галюцинації.
- 8. Проекційний центр чутливості від внутрішніх органів, або аналізатор вісцероцепціі, розташовується в нижній третині постцентральной і предцентральной звивин (поле 43). У корковую частина аналізатора вісцероцепціі надходять аферентні імпульси від гладкої мускулатури і слизових оболонок внутрішніх органів. У корі даній області закінчуються волокна интероцептивного шляху, що відбуваються від нейронів вентролатеральних ядер таламуса, в які інформація надходить по ядерно-таламічна тракт. У проекційному центрі вісцероцепціі аналізуються головним чином больові відчуття від внутрішніх органів і аферентні імпульси від гладкої мускулатури.
- 9. Проекційний центр вестибулярних функцій, безсумнівно, має своє представництво в корі півкуль великого мозку, однак відомості про його локалізації неоднозначні. Прийнято вважати, що проекційний центр вестибулярних функцій розташовується в області середньої та нижньої скроневих звивин (поля 20, 21). Певне ставлення до вестибулярному аналізатору мають також прилеглі відділи тім'яної та лобової часток. У корі проекційного центру вестибулярних функцій закінчуються волокна, що походять від нейронів серединних ядер таламуса. Поразки зазначених кіркових центрів проявляються спонтанним запамороченням, відчуттям нестійкості, почуттям провалювання, відчуттям руху навколишніх предметів і деформації їх контурів.
Завершуючи розгляд проекційних центрів, слід зазначити, що коркові аналізатори загальної чутливості отримують афферентную інформацію з протилежного боку тіла, тому поразка центрів супроводжується розладами певних видів чутливості тільки на протилежній стороні тіла. Коркові аналізатори спеціальних видів чутливості (слуховий, зорової, нюхової, смакової, вестибулярної) пов'язані з рецепторами відповідних органів своєї і протилежної сторін, тому повне випадання функцій даних аналізаторів спостерігається тільки при ураженні відповідних зон кори півкуль великого мозку з обох сторін.
Асоціативні нервові центри. Ці центри формуються пізніше, ніж проекційні, причому терміни кортікалізаціі, тобто дозрівання кори головного мозку, в даних центрах неоднакові. Асоціативні центри відповідають за розумові процеси, пам'ять і реалізацію словесної функції.
- 1. Асоціативний центр"стереогноза", або ядро шкірного аналізатора (центр впізнавання предметів навпомацки). Цей центр розташовується у верхній тім'яної часточці (поле 7). Він двосторонній: в правій півкулі - для лівої кисті, у лівому - для правої. Центр "стереогноза" пов'язаний з проекційним центром загальної чутливості (постцентральна звивина), з якого нервові волокна проводять імпульси больової, температурної, тактильної і пропріоцептивної чутливості. Вступники імпульси в асоціативному кірковій центрі аналізуються і синтезуються, в результаті чого відбувається впізнавання раніше зустрічалися предметів. Протягом усього життя центр "стереогноза" постійно розвивається і вдосконалюється. При ураженні верхньої тім'яної часточки хворі втрачають здатність із закритими очима створювати спільний цілісне уявлення про предмет, тобто не можуть отримати цей предмет на дотик. Окремі властивості предметів, такі як форма, обсяг, температура, щільність, маса, визначаються правильно.
- 2. Асоціативний центр "ПРАКС", або аналізатор цілеспрямованих звичних рухів. Даний центр розташовується в нижній тім'яній часточці в корі надкраевой звивини (поле 40), у правшів - в лівій півкулі великого мозку, у лівш - у правому. У деяких людей центр "ПРАКС" формується в обох півкулях, такі люди в однаковій мірі володіють правою і лівою руками і називаються амбидекстрами.
Центр "ПРАКС" розвивається в результаті багаторазового повторення складних цілеспрямованих дій. У результаті закріплення тимчасових зв'язків формуються звичні навички, наприклад робота на друкарській машинці, гра на роялі, виконання хірургічних маніпуляцій і т.д. У міру накопичення життєвого досвіду центр ПРАКС постійно вдосконалюється. Кора в області надкраевой звивини має зв'язки з задньої і передньої центральними звивинами.
Після здійснення синтетичної та аналітичної діяльності з центру "ПРАКС" інформація надходить в прецентральной звивини до пірамідних нейронам, звідки по корково-спинномозговому шляху досягає рухових ядер передніх рогів спинного мозку.
3. Асоціативний центр зору, або аналізатор зорової пам'яті, розташовується на верхньолатеральну поверхні потиличної частки (поля 18-19), у правшів - у лівій півкулі, у шульг - у правому. У ньому забезпечується запам'ятовування предметів за їх формою, зовнішнім виглядом, кольором. Вважають, що нейрони поля 18 забезпечують зорову пам'ять, а нейрони поля 19 - орієнтацію в незвичній обстановці. Поля 18 і 19 мають численні асоціативні зв'язки з іншими корковими центрами, завдяки чому відбувається интегративное зорове сприйняття.
При ураженні центру зорової пам'яті розвивається зорова агнозія. Частіше спостерігається часткова агнозія (нс дізнається знайомих, свій будинок, себе у дзеркалі). При ураженні поля 19 відзначається викривлене сприйняття предметів, хворий не дізнається знайомих предметів, але він їх бачить, обходить перешкоди.
Нервовій системі людини притаманні специфічні центри. Це центри другої сигнальної системи, що забезпечують здатність спілкування між людьми за допомогою членороздільної людської мови. Людська мова може відтворюватися у вигляді виконання членороздільних звуків ("артикуляція") і зображення письмових знаків ("графіка"). Відповідно в корі головного мозку формуються асоціативні мовні центри - акустичний і оптичний центри мови, центр артикуляції і графічний центр мовлення. Названі асоціативні мовні центри закладаються поблизу відповідних проекційних центрів. Вони розвиваються в певній послідовності, починаючи з перших місяців після народження, і можуть удосконалюватися до глибокої старості. Розглянемо асоціативні мовні центри в порядку їх формування в головному мозку.
4. Асоціативний центр слуху, або акустичний центр промови (центр Верніке), розташований в корі задньої третини верхньої скроневої звивини. Тут закінчуються нервові волокна, що походять від нейронів проекційного центру слуху (середня третина верхньої скроневої звивини). Асоціативний центр слуху починає формуватися на другому-третьому місяці після народження. У міру формування центру дитина починає розрізняти серед оточуючих звуків членороздільно мова, спочатку окремі слова, а потім словосполучення і складні речення.
При ураженні центру Верніке у хворих розвивається сенсорна афазія. Вона проявляється у вигляді втрати здатності розуміти свою і чужу мову, хоча хворий добре чує, реагує на звуки, по йому здається, що оточуючі розмовляють незнайомому йому мовою. Відсутність слухового контролю за власною мовою призводить до порушення побудови речень, мова стає незрозумілою, насиченою безглуздими словами і звуками. При ураженні центру Верніке, оскільки він має пряме відношення до речеобразования, страждає не тільки розуміння слів, але і їх вимову.
5. Асоціативний руховий центр мовлення (речедвігательний), або центр артикуляції мови (центр Брока), розташований в корі задньої третини нижньої лобової звивини (поле 44) в безпосередній близькості від проекційного центру рухових функцій (предцентральная звивина). Речедвігательний центр починає формуватися на третьому місяці після народження. Він односторонній - у правшів він розвивається в лівій півкулі, у шульг - у правому. Інформація з речедвигательного центру надходить у предцентральна звивину і далі по корково-ядерного шляху - до м'язів язика, гортані, глотки, м'язам голови і шиї.
При ураженні речедвигательного центру виникає моторна афазія (втрата мови). При частковому ураженні мова може бути уповільнена, утруднена, скандирована, нескладна, нерідко характеризується лише окремими звуками. Йдеться оточуючих хворі розуміють.
6. Асоціативний оптичний центр мовлення, або зоровий аналізатор письмової мови (центр лексем, або центр Дежерина), знаходиться в кутовій звивині (поле 39). До нейронам оптичного центру мови надходять зорові імпульси від нейронів проекційного центру зору (поля 17). У центрі "Лекс" відбувається аналіз зорової інформації про літери, цифрах, знаках, буквеному складі слів і розумінні їх сенсу. Центр формується з трирічного віку, коли дитина починає дізнаватися букви, цифри і оцінювати їх звукове значення.
При ураженні центру "Лекс" настає алексія (розлад читання). Хворий бачить букви, але не розуміє їх сенсу і, отже, не може прочитати текст.
7. Асоціативний центр письмових знаків, або руховий аналізатор письмових знаків (центр графин), розташовується в задньому відділі середньої лобової звивини (поле 8) поряд з предцентральной звивиною. Центр "графин" починає формуватися на п'ятому-шостому році життя. У цей центр надходить інформація з центру "ПРАКС", призначена для забезпечення тонких, точних рухів руки, необхідних для написання букв, цифр, для малювання. Від нейронів центру "графин" аксони направляються в середню частину предцентральной звивини. Після перемикання інформація по корково-спинномозговому шляху направляється до м'язів верхньої кінцівки. При ураженні центру "графин" втрачається здатність написання окремих букв, виникає "аграфия".
Таким чином, мовні центри мають односторонню локалізацію в корі півкуль великого мозку. У правшів вони розташовуються в лівій півкулі, у шульг - у правому. Слід зазначити, що асоціативні мовні центри розвиваються впродовж усього життя.
8. Асоціативний центр сукупної повороту голови і очей (кортикальний центр погляду) розташовується в середній лобової звивині (поле 9) кпереди від рухового аналізатора письмових знаків (центр графин). Він здійснює регуляцію сукупної повороту голови і очей в протилежну сторону за рахунок імпульсів, що у проекційний центр рухових функцій (предцентральная звивина) від пропріоцепторів м'язів очних яблук. Крім того, в цей центр надходять імпульси від проекційного центру зору (кора в області шпорної борозни - поле 17), що походять від нейронів сітківки ока.
Читайте также:
