
Что такое цикл кори
Ткани и органы. Мышцы
Метаболическая регуляция мышечного сокращения
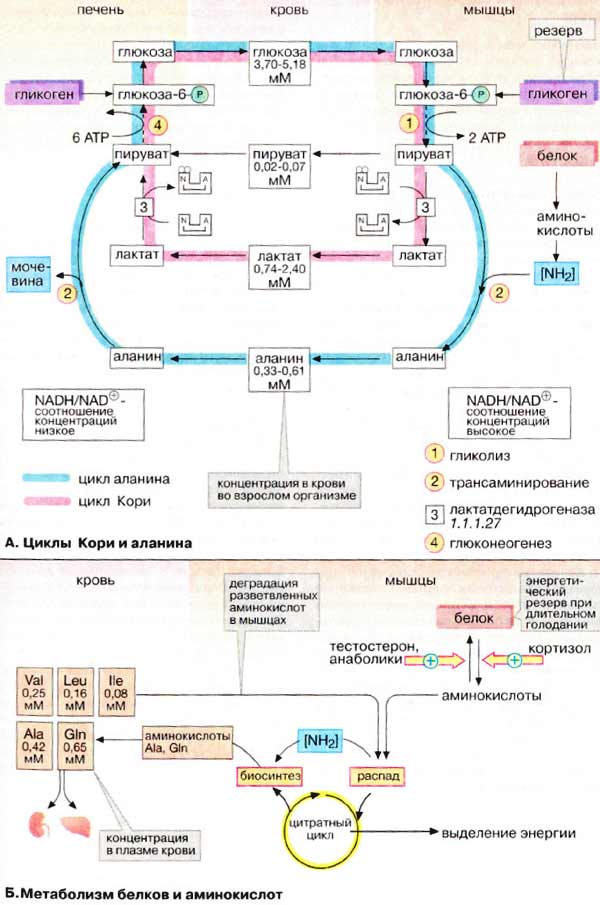
А. Циклы Кори и аланина
В клетках, не содержащих митохондрий (например, в эритроцитах), или в тканях при недостаточном снабжении кислородом (например, в активно работающих мышцах) АТФ (АТР) синтезируется за счет процесса превращений глюкозы в лактат, т. е. за счет процесса брожения (= анаэробного гликолиза , см. рис. 153). Лактат переносится кровью в печень, где в процессе глюконеогенеза с затратой АТФ (см. рис. 157) вновь конвертируется в глюкозу ( цикл Кори ).
При интенсивной работе мышцы максимально активируется гликолиз. Продукт гликолиза, пировиноградная кислота (пируват) накапливается в цитоплазме и недостаточно быстро поступает в митохондрии, если они из-за недостатка кислорода не готовы к окислению пирувата. В анаэробных условиях пируват в реакции, катализируемой лактатдегидрогеназой (см. рис. 103) (заключительный этап гликолиза) восстанавливается до лактата. Одновременно НАДН (NADH), кофермент лактатдегидрогеназы, окисляется до НАД + (NAD + ), который вновь используется на окислительном этапе гликолитического пути. Этой реакции способствует относительно высокое отношение НАДН/НАД + в мышечной ткани. Лактат диффундирует в кровь и поступает в печень, где конвертируется в глюкозу. Таким образом, образование лактата временно заменяет аэробный метаболизм глюкозы и частично переносит этот процесс из мышц в печень.
Обратная связь, подобная циклу Кори, существует в цикле аланина , в котором также участвует пируват . Цикл аланина берет начало с протеолиза белков. Образующиеся аминокислоты в результате трансаминирования в присутствии ферментов превращаются в α-кетокислоты (на схеме не приведено, см. рис. 181), которые в основном включаются в цикл трикарбоновых кислот (цитратный цикл) (см. рис. 183).Одновременно в реакции, катализируемой аланинтрансаминазой, аминогруппы из разных аминокислот переносятся на имеющийся субстрат, пируват. Образующийся аланин поступает в кровь и переносится в печень. Таким образом, цикл аланина служит каналом передачи азота и предшественников глюкозы в печень, которая является местом синтеза конечных продуктов азотистого обмена, например мочевины (см. рис. 185).
Следует напомнить, что при анаэробном гликолизе образуются кислоты, которые, не принимая участие в последующем обмене, существуют в форме анионов. Поэтому при интенсивном анаэробном гликолизе рН мышечной клетки может понизиться настолько, что сокращение станет невозможным. Обычно этого не происходит благодаря быстрому выходу кислых метаболитов (лактата и пирувата) в кровь, которая также может оказаться закисленной (метаболический ацидоз).
Б. Метаболизм белков и аминокислот
Скелетные мышцы активно участвуют в метаболизме аминокислот. Это наиболее важный участок деградации разветвленных аминокислот (Val и Ile, см. с. 402). Ряд других аминокислот также деградируются преимущественно в мышцах. Одновременно идет ресинтез и высвобождение в кровь аланина и глутамина . Эти аминокислоты служат переносчиками азота, образующегося при расщеплении белков, в печень (цикл аланина) и почки (см. рис. 319).
При голодании мышечные белки служат энергетическим резервом организма. Они гидролизуются до аминокислот, которые поступают в печень. Здесь углеродный скелет аминокислот конвертируется в промежуточные продукты цитратного цикла, в том числе в ацетоацетил-КоА и ацетил-КоА (см. рис. 183). Эти амфиболические соединения окисляются в цикле трикарбоновых кислот или включаются в процесс глюконеогенеза.
Синтез и расщепление мышечных белков контролируются гормонами. Тестостерон и синтетические анаболики стимулируют биосинтез белка; напротив, кортизол подавляет образование мышечных белков.
Белки актин и миозин содержат остатки гистидина, метилированного на стадии посттрансляционной модификации. При расщеплении этих белков образуется 3-метилгистидин, который дальше не разрушается. Количество метилгистидина в моче служит мерой деградации мышечных белков.
Цикл Кори (глюкозо-лактатный цикл) открыла чешская ученая, лауреат Нобелевской премии Тереза Кори. Он представляет собой биохимический транспорт лактата из мышц в печень и дальнейший синтез глюкозы из лактата, катализируемый ферментами глюконеогенеза:
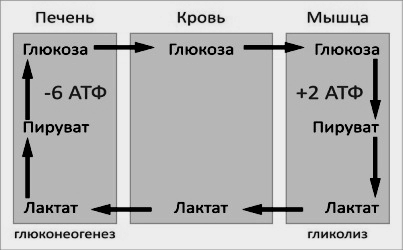
При интенсивной мышечной работе и в условиях отсутствия или недостаточного числа митохондрий (например, в эритроцитах) глюкоза подвергается анаэробному гликолизу с образованием лактата. При накоплении лактата в мышцах возникает лактат-ацидоз, раздражаются чувствительные нервные окончания, что вызывает боль в мышцах.
Лактат переносится кровью в печень и превращается в пируват, а затем в глюкозу (глюконеогенез), которая с током крови может возвращаться в работающую мышцу.
Направление лактатдегидрогеназной реакции в работающих мышцах и печени обусловлено отношением концентраций восстановленной и окисленной форм НАД + : отношение НАД + /НАДН в сокращающейся мышце больше, чем в печени.
7.7. Спиртовое брожение
Спиртовое брожение осуществляется дрожжеподобными организмами, а также некоторыми плесневыми грибками:
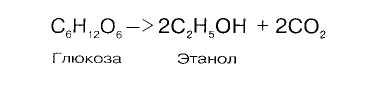
Механизм реакции близок к гликолизу. Расхождение начинается после этапа образования пирувата. При гликолизе пируват при участии фермента ЛДГ и кофермента НАДН восстанавливается в лактат. При спиртовом брожении пируват подвергается декарбоксилированию, в результате образуется ацетальдегид, а затем при восстановлении его - этанол:
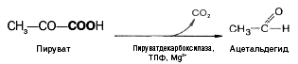

При молочнокислом брожении ПВК не декарбоксилируется, а, как и при гликолизе в животных тканях, восстанавливается при участии ЛДГ за счет водорода НАДН.
7.8. Пентозофосфатный путь превращения глюкозы
Пентозофосфатный путь – альтернативный путь окисления глюкозы. Его функции:
- поставляет клеткам кофермент НАДФН, использующийся как донор водорода в реакциях восстановления;
- обеспечивает клетки пентозофосфатами для синтеза нуклеотидов и нуклеиновых кислот.
Пентозофосфатный путь не приводит к синтезу АТФ.
Ферменты пути локализованы в цитозоле.
В пентозофосфатном пути превращения глюкозы выделяют окислительный и неокислительный пути образования пентоз.
Окислительный путь включает две реакции дегидрирования. Коферментом дегидрогеназ является НАДФ + , восстанавливающийся в НАДФН. Пентозы образуются при окислительном декарбоксилировании.
Неокислительный путь включает реакции переноса 2- и 3-х углеродный фрагментов с одной молекулы на другую. Этот путь служит для синтеза пентоз. Процесс обратим, и из пентоз могут образовываться гексозы.
Пентозофосфатный путь образования пентоз протекает в печени, жировой ткани, молочной железе, коре надпочечников, эритроцитах.
1). Дегидрирование глюкозо-6-фосфата при участии глюкозо-6-фос-фатдегидрогеназы и кофермента НАДФ + с образованием 6-фосфоглюконо--лактона и НАДФН:
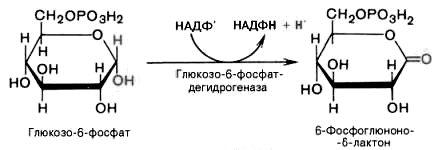
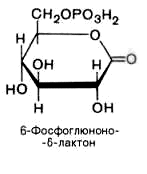
2). 6-фосфоглюконо--лактон нестабилен и гидролизуется с образованием 6-фосфоглюконата (фермент – 6-фосфоглюконолактоназа):
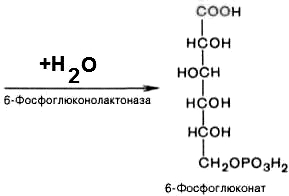
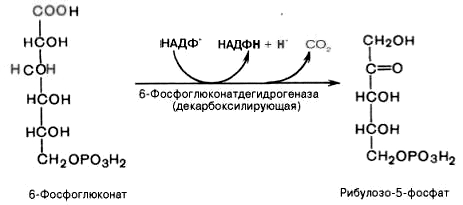
3). Дегидрирование и декарбоксилирование 6-фосфоглюконата с образованием рибулозо-5-фосфата (пентоза) и НАДФН при участии декарбоксилирующей 6-фосфоглюконатдегидрогеназы:
4). Под действием эпимеразы из рибулозо-5-фосфата образуется ксилулозо-5-фосфат (пентоза). Под влиянием изомеразы рибулозо-5-фосфат превращается в рибозо-5-фосфат (пентоза). Между формами пентозофосфатов устанавливается равновесие:
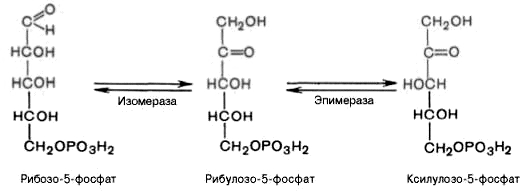
На этом этапе пентозофосфатный путь может быть завершен. При других условиях наступает неокислительная стадия пентозофосфатного цикла, протекающая в анаэробных условиях. Она заключается в переносе двух- и трехуглеродных фрагментов от одной молекулы к другой. При этом образуются вещества, характерные для гликолиза (фруктозо-6-фосфат, фруктозо-1,6-бисфосфат, фосфотриозы), и вещества, специфические для пентозофосфатного пути (седогептулозо-7-фосфат, пентозо-5-фосфаты, эритрозо-4-фосфат).
Шесть молекул глюкозо-6-фосфата в пентозофосфатном цикле образуют 6 молекул рибулозо-5-фосфата и 6 молекул СО2. Из 6 молекул рибулозо-5-фосфата регенерируется 5 молекул глюкозо-6-фосфата:

Промежуточные продукты цикла (фруктозо-6-фосфат и глицеральдегид-3-фосфат) включаются в гликолиз.
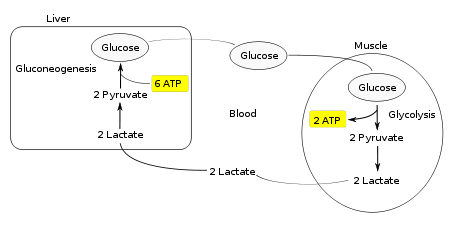
Цикл Кори (также известный как цикл молочной кислоты), названный в честь его открывателей, Carl Ferdinand Cori и Гертите Cori , относится к метаболическому пути , в котором лактат , полученный путь анаэробного гликолиза в мышцах перемещаются в печень и превращается в глюкозу, который затем возвращается к мышцам и метаболизируется обратно в лактат.
содержание

Мышечная деятельность требует АТФ , которая предоставляется пробоем гликогена в скелетных мышцах . Пробой гликоген, процесс , известный как гликогенолиз , высвобождает глюкозу в виде глюкозо-1-фосфата (G-1-P). G-1-Р превращают в G-6-P с помощью фермента фосфоглюкомутазы. G-6-P легко подается в гликолиза , (или может войти в пентозофосфатного пути , если концентрация G-6-P является высоким) процесс , который обеспечивает АТФ в мышечных клетках в качестве источника энергии. Во время мышечной активности, магазин АТФ должен постоянно пополняться. Когда подача кислорода достаточна, эта энергия поступает от кормления пируват , один продукт гликолиза, в цикле Кребса .
Когда подача кислорода недостаточно, как правило , во время интенсивной мышечной активности, энергия должна быть выпущена через анаэробный метаболизм . Брожения Молочная кислота превращает пирувата в лактат посредством лактатдегидрогеназы . Самое главное, что брожение регенерирует NAD + , поддерживая NAD + концентрацию , так что могут возникнуть дополнительные реакции гликолиза. Стадия ферментации окисляет NADH , полученный путем гликолиза обратно в NAD + , передача двух электронов от NADH , чтобы уменьшить пирувата в лактат. (См основных статей по гликолиза и брожения для деталей.)
Вместо того , чтобы накапливать внутри мышечных клеток, лактат получают путем анаэробной ферментации поглощается в печени . Это инициирует вторую половину цикла Кори. В печени глюконеогенез происходит. С точки зрения интуитивной, глюконеогенез меняет как гликолиз и ферментации путем преобразования лактата сначала в пируват, и , наконец , обратно в глюкозу. Глюкозы затем подают к мышцам через кровоток ; она готова подавать в дальнейшие реакции гликолиза. Если мышечная активность прекратилась, глюкоза используется для пополнения запасов гликогена через гликогенез .
В целом, гликолиз часть цикла производит 2 молекулы АТФ на сумму 6 молекул АТФ , потребляемых в части глюконеогенеза. Каждая итерация цикла должен поддерживаться чистым потреблением 4 молекул АТФ. В результате, цикл не может быть устойчивым на неопределенное время . Интенсивное потребление молекул АТФ указывает на то, что цикл Cori сдвигает метаболическую нагрузку с мышц в печень.
Значимость
Важность цикла основывается на предотвращении молочнокислого ацидоза в мышцах в анаэробных условиях. Однако, как правило , прежде чем это произойдет молочная кислота выводится из мышц и в печени.
Цикл также имеет важное значение в производстве АТФ, источник энергии, во время мышечной активности. Функции Cori цикла более эффективно, когда мышечная активность прекратилась. Это позволяет кислородный долг должен быть погашен таким образом, что цикл Кребса и цепь переноса электронов может вырабатывать энергию с максимальной эффективностью.
Цикл Кори является гораздо более важным источником субстрата для глюконеогенеза , чем пищи. Вклад лактата Cori цикла в общем объеме производства глюкозы возрастает с поста длительностью до плато в . В частности, после того, как 12, 20 и 40 часов голодания по добровольцам, глюконеогенез составляет 41%, 71% и 92% продукции глюкозы, но вклад лактата цикла Cori к глюконеогенезу составляет 18%, 35%, и 36 %, соответственно. Остальные производства глюкозы происходит от распада белка, мышечного гликогена, и глицерина из липолиза .
Препарат метформин может вызвать молочный ацидоз у больных с почечной недостаточностью , потому что метформин ингибирует глюконеогенез в печени цикла Кори, в частности митохондриальной цепи комплекса дыхательной 1. Накопление лактата и его субстраты для производства лактата, пируваты и аланина, приводят к избыточному лактату , Обычно избыток лактата будет очищена через почки, но у пациентов с почечной недостаточностью, почки не могут справиться с избыточной молочной кислоты.
Читайте также:
