
Выделение вируса из материала
1. Выделение и идентификация респираторных вирусов
Частота выделения респираторных вирусов зависит от сроков взятия материалов от больных, условий их хранения, доставки и времени первичной обработки. Решающее значение имеет ассортимент и качество клеточных культур и куриных эмбрионов, на которых проводят изоляцию вирусов.
Рекомендуемый состав культур для изоляции вирусов гриппа и ОРЗ:
1) первичная культура клеток почек эмбриона человека (ПЭЧ);
2) перевиваемые культуры клеток МДСК, Неla (или HEp2);
3) фибробласты легкого эмбриона человека (ЛЭЧ).
Материалом для выделения вирусов служат отделяемое носа, зева, конъюнктивы, а также секционный материал (ткани легкого, кусочки бронхов и др.).
Материал отбирают с помощью стерильного тампона, который погружают затем в пробирки с 5,0 мл среды Игла или среды 199 с антибиотиками и немедленно направляют в лабораторию, где тампоны с вирусным материалом отжимают в среду, материал центрифугируют в течение 20 мин при 2000-3000 об/мин и температуре 4°С или используют в неосветленном виде.
Кусочки секционного материала растирают в фарфоровой ступке со стеклянным песком, добавляют 10-кратное количество раствора Хенкса, взвесь ресуспендируют, после чего осветляют центрифугированием. Надосадочную жидкость используют для заражения куриных эмбрионов или тканевых культур.
1.1. Изоляция и идентификация вирусов
Для выделения вирусов гриппа исследуемые материалы после их первичной обработки вводят по 0,1 мл в амниотическую полость и по 0,2 мл в аллантоисную полость 10-дневных куриных эмбрионов.
Заражение проводят в затемненном боксе через боковую поверхность скорлупы, направляя иглу сначала в амниотическую, а затем в аллантоисную полости, предварительно сделав дополнительный прокол в скорлупе над воздушной камерой. Материалом от одного больного заражают 3 эмбриона.
Зараженные эмбрионы инкубируют в термостате 72 часа при 33-34°С. После инкубации эмбрионы помещают на ночь в рефрижератор при 4°С.
Индикацию вируса гриппа осуществляют в реакции гемагглютинации (РГА) путем добавления 0,5 мл 1% суспензии отмытых эритроцитов кур к 0,5 мл вируссодержащего материала. Результаты РГА учитывают после 30-минутного оседания эритроцитов в лунках панелей при комнатной температуре.
При отсутствии агглютинации эритроцитов необходимо провести 3 дополнительных пассажа путем заражения эмбрионов смесью амниотической и аллантоисной жидкостей от предыдущего пассажа. В случае отрицательных результатов РГА после 3-х пассажей исследование материалов прекращают.
Идентификацию вирусов гриппа проводят в РТГА.
Для этого к 0,2 мл последовательных разведений типоспецифических иммунных сывороток добавляют по 0,2 мл рабочей дозы вируса в количестве 4 гемагглютинирующих единиц (ГЕ). Одной ГЕ считают последнее разведение вируса, дающее отчетливую гемагглютинацию. После 1-часового контакта сыворотки с вирусом при комнатной температуре в каждую лунку добавляют по 0,4 мл 1% взвести куриных эритроцитов. Титром сывороток считают ее последнее разведение, блокирующее 4 ГЕ вируса.
Типовую принадлежность вируса определяют по ингибиции гемагглютинации, которая должна быть зарегистрирована в разведении не менее, чем 1:20.
Выделение вирусов гриппа может быть осуществлено на чувствительных культурах клеток, чаще всего МДСК. Для этого в пробирки монослоем, отмытым физиологическим фосфатным буфером или раствором Хэнкса, вносят по 0,2 мл материала от больного. Через 1 ч адсорбции при 38 °С в культуры добавляют по 1,5 мл поддерживающей среды (199, Игла с антибиотиками) и помещают в термостат при 33°С.
Индикацию, типирование и дополнительные пассажи вирусов гриппа проводят как указано выше. Кроме того, инфицированные культуры 1 раз в день микроскопируют для оценки цитопатогенного действия вирусов.
Изоляцию аденовирусов, парагриппозных вирусов, РС-вируса, коронавирусов, вирусов герпеса и др. проводят на клеточных культурах (ПЭЧ, ЛЭЧ и Нер-2). Наиболее чувствительные культуры вносят по 0,2 мл материала от больного. Через 30-60 мин адсорбции вируса на клетках в культуры вносят по 0,8 мл поддерживающей среды Игла с антибиотиками (100 ед/мл пенициллина, 100 мкг/мл стрептомицина) и с 2% инактивированной фетальной сыворотки. Зараженные культуры инкубируют при температуре 36 °С до появления в них проявлений цитопатогенного действия (ЦПД).
Индикацию парагриппозных вирусов проводят по ЦПД и гемадсорбции с 0,4% взвесью эритроцитов морской свинки (20 мин при комнатной температуре), а остальных вирусов
- по характерному цитопатогенному действию.
Если при первичном заражении вирус не был выявлен ни в одном из тестов, материалы дополнительно пассируют еще 2 раза в тех же условиях.
Идентификацию вирусов ОРЗ проводят в чувствительных тканевых культурах с помощью реакции нейтрализации (РН) со специфическими иммунными сыворотками. Вируссодержащую культурную жидкость в дозе 100 ТИД50/0,2 мл в объеме 0,2 мл смешивают с 0,2 мл каждой из типоспецифических иммунных сывороток, взятых в разведении 1:20. Смесь вируса и сыворотки выдерживают 2 часа при комнатной температуре, после чего по 0,2 мл каждой смеси вносят в 2 пробирки с монослоем чувствительных клеток, а затем в пробирки добавляют по 0,8 мл поддерживающей среды. Инфицированные и контрольные культуры клеток инкубируют при температуре 36 °С. Параллельно типоспецифические сыворотки контролируют в отношении их активности к типовым эталонным штаммам.
Учет реакции проводят по ингибиции цитопатогенного действия (для аденовирусов, РС, простого герпеса), гемадсорбции (для парагриппозных вирусов) в день, когда типируемый вирус вызвал выраженное ЦПД клеток в контрольных культурах без сыворотки (при отсутствии дегенерации в контрольных посевах неинфицированных клеток и в контролях диагностических сывороток), в то время как эталонные штаммы полностью нейтрализованы типоспецифическими сыворотками. Тип выделенного штамма определяют по сыворотке, которая полностью предохранила клетки от дегенерации.

ВИРУСОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ , комплекс методов исследования, позволяющих распознать этиологию вирусного заболевания и изучить его возбудителя.
Осн. этапами В. и. являются выделение вируса от больных и павших животных (взятие, консервирование, пересылка и подготовка материала, заражение им животных, куриных эмбрионов, культуры клеток); титрование вирусов для определения их кол-ва в исследуемых материалах; культивирование вирусов на восприимчивых домашних и лабораторных животных, особенно на развивающихся куриных эмбрионах и культурах тканей (гл. обр. первичнотрипсинизированных).
С помощью морфологич. методов выявляют элементарные тельца, внутриклеточные включения (напр., Бабеша — Негри при бешенстве, Боллингера при оспе птиц). Иммунохимич. методы (гл. обр. метод флуоресцирующих антител) позволяют определить специфич. вирусный антиген в заражённых [зараженных] клетках тканевой культуры или органов и тканей инфицир. животных. С помощью серологич. методов проводят видовую и группоспецифич. идентификацию вируса и антител в сыворотках переболевших животных. С этой целью используют реакции нейтрализации, РСК, реакцию гемагглютинации, задержки гемагглютинации, диффузионной преципитации, торможения гемадсорбции. Реакция нейтрализации позволяет улавливать антигенные различия сходных между собой вирусов даже в пределах одноимённой [одноименной] группы. РСК применяют для обнаружения вирусных антигенов в материалах от больных и павших животных, идентификации выделенного вируса, изучения антигенных связей между различными вирусами и определения антител у переболевших животных. Реакции гемагглютинации и задержки гемагглютинации широко применяют в В. и. для диагностич. целей и типирования возбудителей болезни Ньюкасла, гриппа птиц, парагриппа и нек-рых аденовирусов; непрямую реакцию гемагглютинации используют для выявления адено- и миксовирусов, хламидий. Реакцией прицепитации в агаре изучают антигенную структуру вирусов, выявляют антитела в сыворотке и антигены в исследуемом материале. Реакцию гемадсорбции применяют для предварительной индикации и титрования вирусов в культуре клеток, особенно тех, к-рые не вызывают цитопатич. действия. Гемадсорбция наступает раньше проявления цитопатогенного действия, поэтому её [ее] используют как ранний метод предварительной индикации вирусов. Гемадсорбция островковая (адсорбция эритроцитов на отдельных участках монослоя клеток) типична для гриппа свиней, эпидемич. паротита; диффузная (адсорбция эритроцитов на клетках всего монослоя)— для классич. чумы кур, болезни Ньюкасла, афр. чумы свиней и др. Методы очистки и концентрации вирусов используют при изучении физич., химич. и морфологич. свойств вирусов. Фазовые системы, образованные полимерами, используют для концентрации вирусов из больших объёмов [объемов] вируссодержащих жидкостей, выделения нуклеиновых к-т вирусов. Радиобиологич. методы применяют в В. и. для изучения распределения и локализации вирусов (антител) в организме и особенно для изучения процессов онтогенеза вирусов. С помощью электронной микроскопии обнаруживают вирионы в исследуемом материале и специфич. конгломераты вирусов и антител (иммунная электронная микроскопия). Иммуноэлектроосмофорез обусловлен одновременным встречным движением в геле агара антигена и антитела в результате разной величины электрич. заряда с образованием комплекса антиген — антитело в виде линии преципитации и последующей его денатурацией. Метод применяют при изучении антигенной структуры вируса. Изоэлектрич. фокусирование основано на способности белков переходить в инертное (в отношении электрофоретич. подвижности), а затем агрегированное состояние в изоэлектрич. точке. Метод применяют для изучения белков вирусов. При иммуноферментативном методе происходит присоединение фермента к антителам с последующим образованием комплекса фермент — антитело — антиген, к-рый выявляют в клетке с помощью цитохимич. реакции на фермент (пероксидазу, щелочную и кислую фосфотазу, глюкозооксидазу). Метод используют с диагностич. целью для выявления вирусных антигенов в культурах клеток, мазках-отпечатках, криостатных срезах, а также для изучения тонкой структуры антигенов вируса, патогенеза вирусных болезней и др. См. также Вирусоскопия .
Лит.: Тихоненко Т. И., Методические основы биохимии вирусов, М., 1973, с. 152—58; Лабораторная диагностика вирусных и риккетсиозных заболеваний, пер. с англ., ред. Э. Леннет и Н. Шмидт, М., 1974.
Лабораторные исследования при проведении идентификации вирусов и диагностике вирусных инфекций включают следующие этапы: выделение, культивирование, индикация (выявление) и идентификация вирусов.
Вирусы не растут на искусственных питательных средах, а размножаются только внутриклеточно. Крупным достижением было предложение Р. Гудпасчура в 1932 г. использовать для культивирования вирусов куриные эмбрионы. Окончательное решение проблемы культивирования вирусов оказалось возможным лишь после того, как были разработаны основные способы культивирования клеток вне организма.
Использование куриных эмбрионов. Куриные эмбрионы – практически идеальные модели для культивирования некоторых вирусов (например, гриппа и кори). Замкнутая полость эмбриона препятствует проникновению микроорганизмов извне, а также развитию спонтанных вирусных инфекций. Эмбрионы применяют для первичного выделения вирусов из патологического материала; для пассирования и сохранения их, а также для получения необходимых количеств вируса. Некоторые возбудители (например, герпесвирусы) вызывают характерные изменения (по ним можно распознавать заболевание).
Для заражения обычно используют куриные эмбрионы 7–12-дневного возраста. Перед заражением определяют жизнеспособность эмбриона путем овоскопирования (просматривают в проходящем свете). Живые эмбрионы при овоскопировании проявляют двигательную активность, хорошо виден сосудистый рисунок. Простым карандашом очерчивают границы воздушной камеры.
Куриные эмбрионы заражают вируссодержащим материалом в асептических условиях стерильными инструментами, предварительно обработав скорлупу над воздушным пространством йодом и спиртом. Заражение проводят на хорион-аллантоисную оболочку, в амниотическую или аллантоисную полость, либо в желточный мешок (рисунок 18). Выбор метода заражения зависит от биологических свойств вируса.

Рисунок 18 – Схематическое изображение развивающегося куриного эмбриона
Культура клеток. Вначале был использован метод переживающих тканей. Он заключался в том, что в колбу, содержащую питательную среду, вносили кусочек ткани. Клетки некоторых тканей в таких условиях могут переживать (но не размножаться) до 30 дней, а в них могут размножаться вирусы. Однако этот способ давал очень небольшой выход вирусов. Необходимо было разработать условия, при которых клетки ткани могли бы свободно размножаться.
Для получения культур клеток необходимо было решить четыре главных задачи:
получить в необходимом количестве свободные (т. е. изолированные друг от друга) клетки;
создать такие питательные среды и условия, в которых клетки могли бы активно размножаться;
обеспечить условия, при которых в культурах клеток не могли бы размножаться бактерии;
определить методы, с помощью которых можно было бы распознавать рост вируса в культуре клеток и идентифицировать его.
Для выделения изолированных (разобщенных), но жизнеспособных клеток из разрушенных тканей, стали использовать обработку их слабым раствором трипсина, разрушающего межклеточные мостики. Для культивирования клеток были предложены различные среды, содержащие все необходимые для размножения клеток питательные вещества (аминокислоты, основания, витамины и другие), минеральные соли, имеющие оптимальную рН и т. д. К питательным средам добавляли индикатор, по изменению цвета которого можно было судить о метаболизме клеток и их размножении. Было установлено, что в качестве основы, на которой клетки размножаются и образуют монослой, может быть использовано хорошо обработанное стекло пробирок и колб. Для подавления возможного роста бактерий вируссодержащий материал перед посевом его в культуры клеток стали обрабатывать антибиотиками.
В 1949 г. Дж. Эндерс, Т. Веллер и Ф. Роббинс показали, что вирус полиомиелита хорошо размножается в первично-трипсинизированных культурах клеток, полученных из почек обезьян. Основной недостаток первично-трипсинизированных клеток заключается в том, что после нескольких пересевов они перестают размножаться. Поэтому предпочтением стали пользоваться культуры таких клеток, которые способны размножаться in vitro бесконечно долго. Такие перевиваемые культуры клеток (клеточные линии характеризуются бессмертием и гетероплоидным кариотипом) получают из опухолевых тканей (HeLa получена из карциномы шейки матки, НЕр-2 – из карциномы гортани; Детройт-6 – из метастаза рака легкого в костный мозг; RН – из опухоли почки человека) или из мутантных клеток с полиплоидным набором хромосом. Однако опухолевые клетки нельзя применять для получения вакцин. Для этих целей используют только культуры таких клеток, которые не содержат никаких контаминантных вирусов и не обладают злокачественностью. Лучше всего этим требованиям отвечают культуры диплоидных клеток.
Полуперевиваемые (диплоидные) культуры клеток – клетки одного генотипа, способные in vitro выдерживать 50–100 пассажей, сохраняя при этом свой исходный диплоидный набор хромосом. Диплоидные линии фибробластов эмбриона человека используются как для диагностики вирусных инфекций, так и при производстве вирусных вакцин. Как оказалось, вирусы могут размножаться не только в культурах клеток, образующих монослой на стекле пробирок, но и в суспензиях живых клеток.
Для обеспечения жизнедеятельности культивируемых клеток необходимы питательные среды. По назначению они делятся на ростовые и поддерживающие. В ростовых питательных средах должно содержаться больше питательных веществ, обеспечивающих активное размножение клеток и формирование монослоя. Поддерживающие среды обеспечивают переживание клеток в уже сформированном монослое в период размножения в них вирусов. вирус заражение культивирование инфекция
Выделение вирусов в культурах клеток. При выделении вирусов из различных инфекционных материалов (кровь, моча, слизистые отделяемые, смывы из органов) применяют культуры клеток, обладающих наибольшей чувствительностью к предполагаемому вирусу. Для заражения используют культуры в пробирках с хорошо развитым монослоем клеток. Перед заражением клеток питательную среду удаляют и в каждую пробирку вносят по 0,1–0,2 мл взвеси исследуемого материала, предварительно обработанного антибиотиками для уничтожения бактерий и грибов. После 30-60 мин контакта вируса с монослоем клеток удаляют избыток материала, в культуру вносят поддерживающую среду и пробы оставляют в термостате до выявления признаков размножения вируса.
Выделение вирусов на лабораторных животных. При невозможности выделить и идентифицировать вирус стандартными методами in vitro инфекционный материал вводят чувствительным к возбудителю животным, и после развития типичного инфекционного процесса проводят повторное заражение чувствительных клеточных культур. Наиболее часто используют мышей, кроликов и обезьян; для выделения некоторых вирусов (например, вирусов Коксаки) заражают мышат-сосунков. Вследствие дороговизны и сложности содержания лабораторных животных, практически повсеместно их вытеснили клеточные культуры. Тем не менее животные модели активно используют для изучения особенностей патогенеза и формирования иммунных реакций при вирусных инфекциях.
Таким образом, для выделения чистых культур вирусов в лабораторных условиях в настоящее время используются следующие живые объекты (биологические модели): 1) культура клеток (тканей, органов); 2) куриные эмбрионы; 3) лабораторные животные.
Индикация вирусов в культурах клеток. Индикатором наличия вируса в зараженных культурах клеток может служить:
1) развитие специфической дегенерации клеток – цитопатическое действие вируса (ЦПД), имеющее три основных типа: крупно- или мелкоклеточная дегенерация; образование многоядерных гигантских клеток (симпластов); развитие очагов клеточной пролиферации, состоящих из нескольких слоев клеток (гроздевидная дегенерация клеток).
Различают два механизма гибели клеток, вызываемой вирусами, – некроз и апоптоз. Некроз происходит из-за необратимых нарушений целостности клеточных мембран, апоптоз – вследствие фрагментации ядерной ДНК под действием клеточной эндонуклеазы.
Цитопатические эффекты оценивают при микроскопии клеточных культур. По степени поражения клеток выделяют вирусы с высокой или умеренной цитопатогенностью:
2) обнаружение внутриклеточных включений, располагающихся в цитоплазме и/или в ядрах пораженных клеток. При ряде вирусных болезней обнаружение телец-включений имеет диагностическое значение. Многие из них настолько патогномоничны, что обнаружение их, стало основным из экспресс методов диагностики бешенства, оспы, ринопневмонии лошадей, аденовирусной инфекции и др..;
3) положительная реакция гемагглютинации (РГА) или гемадсорбции (РГАдс). Некоторые вирусы, в частности, вирус гриппа, обладают особыми рецепторами (гемагглютининами), с помощью которых они адсорбируются на эритроцитах и вызывают их склеивание (гемагглютинацию). Такие вирусы легко обнаруживаются с помощью реакции гемагглютинации или гемадсорбции (эритроциты адсорбируются на инфицированных вирусами клетках культуры тканей);
4) феномен бляшкообразования. Широкое распространение получил предложенный в 1952 г. Р. Дюльбекко метод бляшек (негативных колоний), позволяющий производить количественное определение вирусов. Для выделения вирусов монослой клеток после удаления питательной среды заражают вируссодержащим материалом и покрывают слоем агара, содержащего индикатор нейтральный красный. Чашки (флаконы) инкубируют при 37 °С. Через 48–96 ч выявляются пятна – бляшки. Они имеют диаметр 1–3 мм и выглядят неокрашенными на розовом фоне. Пятна возникают за счет цитопатического действия вируса;
5) цветная реакция Солка. О росте вирусов в клетках можно судить с помощью индикатора, добавляемого к питательной среде. Если клетки активно осуществляют метаболизм, рН среды сдвигается в кислую сторону, и среда окрашивается в желтый цвет. В случае размножения вируса клетки погибают, рН среды мало меняется, и она сохраняет первоначальный (малиновый) цвет или (при нейтральной рН) приобретает оранжевый;
6) реакция интерференции (используется при отсутствии ЦПД, гемагглютинации и гемадсорбции): исследуемая культура повторно заражается вирусом, вызывающим ЦПД. В положительном случае ЦПД будет отсутствовать (реакция интерференции положительна). Если в исследуемом материале вируса не было, наблюдается ЦПД.
Кроме того, для обнаружения вируса в культурах клеток могут быть использованы различные серологические реакции.
Индикация вирусов на лабораторных животных. Индикация вируса основана на обнаружении у животных признаков инфекционного заболевания, регистрации их гибели, изучении характера патоморфологических и патогистологических изменений в тканях и органах, выявлении положительной реакции гемагглютинации.
Вирусы являются внутриклеточными инфекционными агентами. Весь репликативный цикл вируса осуществляется с исполь зованием метаболических и генетических ресурсов клеток. Поэтому патогенез вирусных инфекций, в первую очередь, следует рассматривать на молекулярном и клеточном уровнях (В.И. Покровский, О.И. Киселев, 2002). Вместе с тем, инфекционный процесс, вызванный вирусами, развивается в пределах того или иного органа или ткани, так как большинство вирусов обладают достаточно высокой органной или тканевой тропностью. Поэтому характер развития внутритканевых процес сов при вирусных инфекциях, с одной стороны, определяется, как правило, цитопатическим действием вируса на клетки данной ткани и органа, а с другой стороны, реакцией внутритканевых и органных систем защиты от вирусной инфекции. Последние не редко носят деструктивный характер и усугубляют течение забо левания и его последствия. Поэтому понимание патогенеза вирусных инфекций представляется исключительно важным для создания рациональных схем лечения, основанных на гармоничном сочетании симптоматического лечения и противовирусной химиотерапии.
Стадии репликативного цикла вируса гриппа представлены на рис. 1. Такой же репликативный цикл свойствен большинству РНК- и ДНК-содержащих вирусов, кроме тех, кото рые обладают особыми механизмами хронизации процесса, латенции и способностью к интеграции в клеточный геном (например, ВИЧ, гепатит С, вирус папилломы и.т.). Ключевыми стадиями репликативного цикла вируса являются проникновение в клетки, декапсидация, активация процессов транскрипции и трансляции, собственно репликация вирусного генома и созревание зрелых вирусных частиц с выходом нового потомства инфекционного вируса.
Проникновение вируса в клетки - одна из ключевых стадий патогенеза инфекционного процесса. Для большинства вирусов степень цитопатического действия вируса прямо зависит от множественности заражения. В естественных условиях инфицирова ние клеток вирусами не происходит обычно с высокой множе ственностью. Исключение составляют респираторные вирусы, которые передаются воздушно-капельным путем с высокой инфицирующей дозой. ВИЧ и гепатиты, наоборот, эффективно передаются с кровью в самых незначительных трудно определяе мых титрах. Такие вирусы имеют особые свойства, определяющие их контагиозность.
Для проникновения вируса в клетку, необходимо: высокое сродство к вирусоспецифическому рецептору; множественность рецепторов; эффективность слияния вируса с клетками при инфицировании; способность к образованию синцития, к образованию гигантских синпластов, состоящих из множества слившихся клеток, в которых вирусный нуклеопротеид без эндоцитоза беспрепятственно переходит от одной зараженной клетки к множеству других через цитоплазму.
Такими свойствами из респираторных вирусов обладает респираторно-синцитиальный вирус, а среди ретровирусов – вирус иммунодефицита человека.
Стадии жизненного цикла вируса гриппа и соответствующие ингибиторы репродукции в известной степени могут быть экстраполированы на многие другие вирусы, за исключением тех, жизненный цикл которых включает интеграцию в клеточный геном. В процессе репликации вирусов обычно происходит селективная блокада трансляции клеточных мРНК и активация экспресси клеточных генов, кодирующих провоспапительные лимфокины. Многократные раунды репликации вирусного генома приводят к истощению энер гетических ресурсов клеток и пула предшественников азотистых оснований. Повышенное содержание провоспалительных лимфокинов приводит к развитию воспалительной реакции и локально му генерированию свободных радикалов, оказывающих, в свою очередь, повреждающее действие на клетки и ткани через перекисное окисление липидов клеточных мембран, инактивацию жизненно важных ферментов. Поэтому одним из важнейших факторов патогенности вирусов является их репликативный потенциал: чем активнее происходит репликация, тем сильнее цитопатическое действие вирусов на клетки и пораженную ткань (орган).
Значительный вклад в цитопатическое действие вирусов вносит репродукция вирусов в митохондриях, что сопровождается нарушением функций этих органоидов, падением продукции АТФ и индукцией опосредованного митохондриями пути апоптоза.
Впервые возможность репродукции вирусов в митохондриях была доказана выдающимся отечественным вирусологом В.М. Ждановым. Узкий тропизм вирусов, в известной степени, спасителен для организма, так как инфекционный процесс ограничен одним органом.
Это обстоятельство позволяет, проводит рациональную симптоматическую терапию, направленную на защиту данной ткани или органа и ориентировать противовирусные средства в данный орган, например, легкие.
Возбудители гриппа относятся к семейству ортомиксовирусов (Orthomyxo-viridae) и являются пневмотропными вирусами. Схематическое строение вируса гриппа представлено на рис. 2.1. Геном вируса складывается из 8-ми фрагментов однонитчатой РНК, которые кодируют
10 вирусных белков. Фрагменты РНК имеют общую белковую оболочку, соединяющую их, образуя антигенно-стабильный рибонуклеопротеид (S-антиген), который определяет принадлежность вируса к серотипу А, В или С. Снаружи вирус покрыт двойным липидным слоем с внутренней стороны которого находится слой мембранного белка.
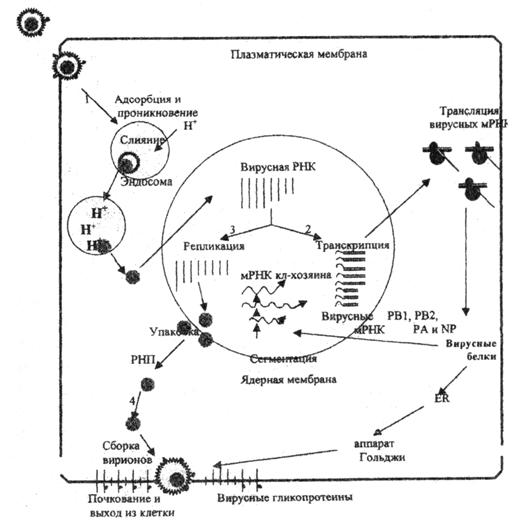
Рис. 1. Репликативный цикл вируса гриппа (В.И.Покровский, О.И.Киселев, 2002)
Представлены 4 основные фазы репликативного цикла вируса гриппа: 1 – адсорбция и проникновение вируса в клетку хозяина; 2 – транскрипция вирусной РНК и трансляция вирусных белков; 3 – репликация вирусной РНК и 4 – самосборка вирионов и последующий выход их из клетки.
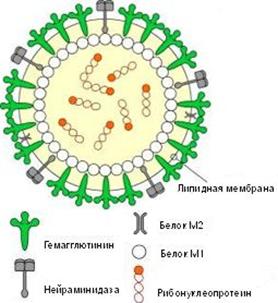
Рис. 2. Схематическое строение вируса гриппа
Гемаглютинин и нейраминидаза являются факторами агрессии вируса гриппа. Интенсивность интоксикации при гриппе определяется свойствами гемаглютинина, а нейраминидаза обладает выраженным иммунодепресивным действием. Оба поверхностных антигена характеризуются выраженной способностью к изменчивости, в результате чего появляются новые антигенные варианты вируса. Гемаглютинины 1, 2, 3 типов и нейраминидазы 1, 2 типов содержат вирусы, которые поражают человека. Другие антигены характерные для вирусов гриппа животных (свиней, собак, лошадей, многих видов птиц и др.).
Вирусы гриппа А. Эти возбудители более вирулентны и контагиозны, чем вирусы гриппа В и С. Это обусловлено тем, что вирус А содержит 2 типа нейраминидазы (N1, N2) и 4 –гемаглютинина (H0, H1, H2, H3); вирус В – 1 и 1 соответственно, поэтому он имеет лишь антигенные варианты внутри одного серотипа; вирус С содержит только гемаглютинин и не содержитнейраминидазу, но он имеет рецептор, который разрушает энзим. Вирусу С не свойственная изменчивость.
Одновременно циркулируют и имеют эпидемическое распространение 2 подтипа вируса гриппа А H3N2 и H1N1. На сегодняшний день во всем мире доминирует вариант А/Сидней/05/97, А/Берн/07/95 и А/Пекин/262/95. Характерной чертой современных вирусов гриппа А (H3N2) является изменение ряда их биологических свойств: тяжело размножаются в куриных эмбрионах, не агглютинируют эритроциты кур, взаимодействуют только с эритроцитами человека или морской свинки, имеют низкую иммуногенную активность.
Вирусы гриппа В выделяются только от людей. По вирулентности и эпидемиологической значимости они уступают вирусам гриппа А. По антигенным свойствам гемаглютинин инейраминидазу вируса В можно разделить на 5 подтипов. Их изменчивость носит более постепенный и медленный характер, что объясняет эпидемиологические особенности вируса гриппа В (эпидемии возникали 1 раз в 3-4 года до 1988 г.). В мире сейчас циркулируют 2 варианта вируса гриппа В: В/Пекин/184/93 (Европа, Америка, Африка, Австралия) и В/Виктория/2/87 (Юго-Восточная Азия). Характерной чертой современных вирусов гриппа А и В является их высокая чувствительность к ингибиторам гемаглютинирующей и инфекционной активности, которые содержатся в сыворотке крови нормальных животных.
Вирусы гриппа С . Вирусы гриппа С, в отличие от вирусов А и В, не вызывают эпидемий, но они могут быть причиной вспышек в организованных детских коллективах. Подъемы заболеваемости гриппом С нередко предшествуют или сопровождают эпидемии гриппа А и В. У детей раннего возраста и более старших возрастных группах он приводит к заболеваниям в легкой ибессимптомной форме.
Для вирусов гриппа С характерна значительно большая стабильность антигенных и биологических свойств. По ряду биологических характеристик эти возбудители отличаются от других представителей семейства ортомиксовирусов. Им присуща низкая репродуктивная активность в разных клеточных системах и наличие других, чем у вирусов гриппа А и В, рецепторов на поверхности эритроцитов. Рецептор-деструктирующая активность связана не с нейраминидазой, как у вирусов гриппа А и В, а с ферментом нейраминат-0-ацетилестеразою.
Сложность этиологии усиливают факты выделения от людей вирусов, которые не свойственные человеку. Гены вируса гриппа (H5N1), выделенные в 1997 году от человека в Гонконге, оказались подобными генам птичьего вируса, который преодолел межвидовой барьер. Оказалось, что человеческие и птичьи вирусы гриппа были реасортантами (рекомбинантные штаммы), которые получили внутренние гены РВ1 и РВ2 от перепелиного вируса гриппа А (H9N2) – штаммы А/перепел/Гонконг/61/97.
Вирусы гриппа малоустойчивы во внешней среде, в течение нескольких часов при комнатной температуре разрушаются. Под действием дезинфицирующих растворов (спирта, формалина, сулемы, кислот, щелочей) они быстро погибают. Нагревание до 50-60°С инактивирует вирусы в течение нескольких минут, в замороженном состоянии при температуре -70° С они сохраняются годами, не теряя инфекционных свойств, быстро погибают под воздействием ультрафиолетового излучения. Оптимум размножения вируса происходит при +37°С в слабощелочной среде. Как и другие вирусы, возбудитель гриппа отнюдь не чувствительный к антибиотикам и сульфаниламидам.
Вирус гриппа не растет на обычных питательных средах. Его можно выделить из материала, который получен от больного в первые дни болезни (смывы из носоглотки, мокроты), путем заражения культур или клеток куриных эмбрионов.
Традиционно считалось, что человеческий организм освобождается от возбудителя в ближайшие дни после выздоровления. Однако постепенно накапливались косвенные свидетельства, которые опровергают эту точку зрения. И в 1985 г. были получены прямые доказательства формирования длительной персистенции вируса гриппа. У одного человека удалось многократно выделять вирус гриппа в течение 9-ти месяцев и дольше. Возможность пожизненной персистенции вируса в организме человека продолжает изучаться.
1. Грипп и другие респираторные вирусные инфекции / под ред. О.И. Киселева, И.Г. Мариничева, А.А. Сомининой. – СПб. – 2003.
2. Дриневский В.П., Осидак Л.В., Цыбалова Л.М. Острые респираторные инфекции у детей и подростков // Практическое руководство под редакцией О.И. Киселева. – Санкт-Петербург. – 2003.
3. Железникова Г.Ф., Иванова В.В., Монахова Н.Е. Варианты иммунопатогенеза острых инфекций у детей. – Санкт-Петербург. – 2007. – 254 с.
4. Ершов Ф.И. Грипп и другие ОРВИ // Антивирусные препараты. Справочник. – М., 2006. –С. 226-247.
5. Ершов Ф.И., Романцов М.Г. Антивирусные средства в педиатрии. – М., 2005. – С.159-175.
6. Ершов Ф.И., Киселев О.И. Интерфероны и их индукторы (от молекул до лекарств). – М., 2005. – С. 287-292.
7. Иванова В.В. Острые респираторно-вирусные заболевания // Инфекционные болезни у детей. – М., 2002.
8. Онищенко Г.Г., Киселев О.И., Соминина А.А. Усиление надзора и контроля за гриппом как важнейший элемент подготовки к сезонным эпидемиям и очередной пандемии. – М., – 2004. – С.5-9.
9. Об утверждении стандарта медицинской помощи больным гриппом, вызванным идентифицированным вирусом гриппа (грипп птиц) // Приказ Минздравсоцразвития №460 от 07.06.2006 г.
10. Романцов М.Г., Ершов Ф.И. Часто болеющие дети: Современная фармакотерапия. – М., 2006. – 192 с.
11. Стандартизированные принципы диагностики, лечения и экстренной профилактики гриппа и других острых респираторных инфекций у детей / под ред. О.И. Киселева. – СПб. – 2004. – С. 82-95.
12. Лекарственные средства в фармакотерапии патологии клетки / под редакцией Т.Г. Кожока.-М., 2007.
Читайте также:
