
Внутривидовой и межвидовой обмен генетическим материалом у вирусов
Генетика микробов
Изменчивость и наследственность микроорганизмов является частью общебиологической проблемы изменчивости и наследственности. Наследственность и изменчивость в сущности две стороны одного явления. В природе постоянно наблюдается процесс передачи наследственных свойств организма из поколения в поколение и в то же время идет процесс изменчивости. Наследственность и изменчивость - это два противоречивых и вместе с тем неразрывно связанных между собой процесса. Они основа развития живого мира. По сравнению с другими организмами изменчивость микробов наблюдается чаще и осуществляется легче и быстрее, это объясняется большой быстротой их размножения и пластичностью.
Изменяться могут самые разнообразные свойства микробов - морфологические, ферментативные, антигенные, патогенные и др. Различают ненаследственную и наследственную изменчивость. Ненаследственная изменчивость (модификация) очень часто наблюдается под воздействием различных факторов внешней среды. Она заключается в количественном изменении некоторых свойств микроба, т. е. в ослаблении и утрате или усилении этих свойств. Когда воздействие факторов, вызвавших эти изменения, прекращается, то возникшие измененные признаки также утрачиваются. Наследственная изменчивость необратима, она развивается вследствие перестройки наследственного аппарата микроорганизма в результате непосредственного внешнего или внутреннего воздействия на него или внедрения чужеродного генетического материала (трансформация, конъюгация, рекомбинация и др.).
В микробиологии основоположником учения об изменчивости является Луи Пастер. Он искусственным путем получил необратимое ослабление вирулентности возбудителей сибирской язвы (1881) и бешенства (1885), а также получил способ приготовления живых вакцин для борьбы с этими заболеваниями. Это открытие Пастера имело поворотное значение для борьбы с инфекционными заболеваниями человека и животных. Применение живых вакцин привело к ликвидации многих эпидемий, сведя их до единичных заболеваний. Противосибиреязвенная вакцина получена Пастером путем длительного (12-24 дня) выращивания при повышенной температуре (42°) вирулентной культуры сибиреязвенной палочки.
Путем такой направленной изменчивости микробов в настоящее время получено 20 живых вакцин, из них 14 против вирусных и 6 против бактериальных инфекций. Все время идут работы по созданию новых вакцин и улучшению некоторых из существующих.
Благодаря простоте своего строения вирусы обладают большой способностью к изменчивости. Вирусы растений способны приспособляться к условиям жизни в организмах филогенетически очень отдаленных видов. Так, к вирусу табачной мозаики восприимчивы 236 видов растений 33 различных семейств. Из животных вирусов наибольшей изменчивостью обладает вирус гриппа. Вирусы гриппа, выделенные во время отдельных эпидемий в различных странах, отличаются от основных типов А и В по антигенному строению, морфологическим признакам, способности к адаптации и пр. В 1947-1952 гг. в Китае появилась новая разновидность - тип А2, которая вызвала эпидемию гриппа во всем мире. Об изменчивости вирусов говорит тот факт, что наибольшее количество живых вакцин получено из вирусов.
Изменчивость грибов имеет большое практическое значение, так как многие из них широко используются в различных производствах. Были получены расы дрожжей, хорошо развивающиеся при пониженных или повышенных температурах, требуемых в тех или иных производствах. Для интенсификации бродильных процессов получены расы дрожжей, способные использовать углеводы в больших концентрациях, а также приспосабливаться к высокому осмотическому давлению, к продуктам собственной жизнедеятельности, к различным физическим и химическим факторам. Применение этих рас дает большую продукцию. Ярким примером этого является производство антибиотиков.
За последние годы в изучении наследственности микроорганизмов наблюдаются большие достижения, имеющие значение не только для микробиологии, но и для общей генетики, биохимии и других наук. Проведенные генетические исследования приближают нас к реальной возможности влияния на процессы наследственности и изменчивости микробов.
Исходными для понимания явлений наследственности служат данные о химическом составе хромосом. Хромосомы высших организмов являются нуклеопротеидами, генетическим материалом в которых служит ДНК. У бактерий и фагов хромосомами является непосредственно ДНК, на нити которой в линейном порядке на определенных местах расположены определенные наследственные признаки. Количество хромосом в клетках различных организмов постоянно: у человека 46 хромосом, у кукурузы 10 и т. д. У бактерий весь генетический материал представляет одну хромосому. Это было найдено при помощи генетического метода.
Наследственность неразрывно связана с размножением, а размножение - с делением клеток, при котором происходит саморепродукция хромосом и ДНК. Соматические клетки имеют диплоидный, двойной набор хромосом. Дочерние клетки имеют такое же число хромосом, как и материнская (митоз). В половых клетках содержится вдвое меньше хромосом, так как при образовании половых клеток (мейоз) происходит особое редукционное деление с уменьшением нормального числа хромосом наполовину, например в половых клетках человека 23 хромосомы. Такой набор хромосом называется гаплоидным.
Из микроорганизмов только некоторые грибы и водоросли имеют типичный половой процесс с диплоидным и гаплоидным набором хромосом. Бактерии и фаги генетически ведут себя как организмы с одной гаплоидной хромосомой.
Характерно, что обычные морфологические методы смогли дать только общее описание ядерного вещества, применение же генетического метода исследования раскрыло тонкую структуру бактериальной хромосомы, хотя понятие хромосомы у бактерий носит несколько условный характер. Ген в настоящее время не является чисто умозрительной категорией, понятие гена получило вполне материальное содержание. Ген представляет собой участок (локус) хромосомы, слагающийся из цепи нуклеотидов молекулы ДНК, обладающий специфической функцией. Хромосома - это цепь линейно расположенных генов. Участок молекулы ДНК, составляющий ген, включает от 500 до нескольких тысяч нуклеотидов. Термину "ген" равнозначны: наследственный фактор, генетическая единица, детерминанта и др. Совокупность всех наследственных факторов (генов) в хромосомах организма называется генотипом. Фенотипом является внешний вид организма со всеми внешними и внутренними признаками. Фенотип бактерии есть результат взаимодействия ее генотипа и среды.
Появление в каком-либо организме новых наследственно передаваемых свойств может быть вызвано разными типами изменений в генетическом аппарате. Источником наследственной изменчивости могут быть мутации генов, т. е. изменения в их химическом строении, и структурные мутации хромосом. При этом могут происходить в двойной спирали ДНК следующие изменения: 1) замещение пары оснований, имевшихся в исходной молекуле ДНК, другой парой; 2) выпадение пары оснований из молекулы ДНК (деления); 3) внедрение новой пары оснований в молекулу ДНК; 4) инверсия - поворот нескольких пар оснований на 180°. Таким образом, в основе мутаций лежат молекулярные изменения в хромосоме.
Мутации у бактерий выявляются при наследственных изменениях любого признака микроба. Наиболее легко выявляются и количественно точно учитываются такие признаки: ауксотрофность к аминокислотам, пуринам и пиримидинам, витаминам; чувствительность или устойчивость к антибиотикам или фагам; лизогенность; ферментация углеводов и др. Предложены определенные обозначения мутантов. Так, ауксотрофность аминокислот обозначают начальными буквами или слогами их, например: гистидина - his, триптофана - try и т. д., двойные, тройные ауксотрофы обозначаются his try. Способность ферментировать углеводы, например лактозу - 1ас + , неспособность ферментировать - 1ас - . Чувствительность к стрептомицину - Str-s, резистентность - Str-r, зависимость - Str-d и т. д. Под влиянием мутагенных факторов были получены ауксотрофные мутанты, которые потеряли способность к синтезу важных для микроорганизмов веществ. Так, одни мутанты потеряли способность синтеза тех или иных аминокислот, другие мутанты - синтеза витаминов и т. д. Эти ауксотрофы (минус-варианты) могут расти только на средах, содержащих вещества, синтезировать которые они уже не могут. У них нарушена активность соответствующих ферментов. С помощью таких ауксотрофов удалось проследить нормальный биосинтез многих аминокислот, витаминов, углеводов, азотистых оснований.
Мутации у микробов так же, как и у высших организмов, делятся на спонтанные и индуцированные. Спонтанные мутации возникают при воздействии факторов, являющихся нормальными условиями среды. Индуцированные мутации возникают в результате обработки микробов мутагенными агентами. К последним относятся различные виды радиации (ультрафиолетовые, рентгеновские лучи, быстрые нейтроны, протоны и др.), действие температуры и пр. Спонтанные мутации наблюдаются очень редко. Частота их колеблется от 10 10 до 10 4 , т. е. одна мутантная клетка на 10 тыс. млрд. клеток. Мутагены увеличивают частоту мутаций в 10-100 тысяч раз.
Представление о строении генетического аппарата бактерийной клетки основано на изучении у них механизмов генетического обмена. У бактерий нет такого полового процесса, какой имеется у высших организмов. У бактерий обычно происходит только односторонняя передача части наследственного материала (фрагмента ДНК) от одной бактерийной клетки - донора к другой - реципиенту. При этом в клетке реципиента происходит взаимодействие между генетическим материалом донора и реципиента, которое приводит к образованию дочерних рекомбинантных клеток. Последние клетки сохраняют некоторые признаки реципиента и приобретают новые признаки, полученные от клетки донора. Механизмами такой рекомбинации, перегруппировки признаков в потомстве, т. е. изменчивости, являются трансформация, трансдукция и конъюгация.
Трансформация (превращение, перестройка) заключается в том, что некоторые бактерии при выращивании их в присутствии веществ, извлеченных из клеток родственных им видов или разновидностей, приобретают некоторые свойства последних. Пневмококки - возбудители воспаления легких - имеют несколько разновидностей. Когда к неболезнетворной бескапсульной разновидности прибавили убитые микробные тела болезнетворной разновидности, имеющей капсулу, и ввели эту смесь белым мышам, то белые мыши, вопреки ожиданию, погибли и из крови их была выделена болезнетворная капсульная разновидность (Ф. Гриффитс, 1928). Контрольные мыши, получившие только убитые микробные тела, не заболели. Таким образом, в организме мышей неболезнетворный пневмококк приобрел свойства болезнетворного, хотя и убитого, пневмококка. Это свойство стало наследственно закрепленным. После больших исследований было найдено, что эта наследственная изменчивость была вызвана ДНК убитых микробных тел болезнетворного пневмококка (О. Т. Эвери, 1944). В дальнейшем такие трансформации были получены при помощи одной только ДНК, полученной из микробных тел. Так же хорошо изучена трансформация устойчивости к антибиотикам - пенициллину, стрептомицину. Если ДНК, выделенную из антибиотиков устойчивых бактерий, добавить в культуру чувствительных к антибиотикам бактерий, то в культуре под влиянием ДНК некоторое количество клеток приобретает наследственную устойчивость к данному антибиотику. Активность ДНК, измеряемая в гаммах на 1 мл среды (v=10 -6 г), оказалась чрезвычайно высокой. Показана также возможность и межвидовой трансформации. Установление генетической роли ДНК имеет общебиологическое значение, так как позволило перейти к изучению различных биологических процессов на молекулярном уровне (синтез белка, нуклеиновых кислот, кодирование и др.).
Трансдукция - наследственно закрепленная передача признаков от одной бактерийной клетки (донора) другой клетке (реципиенту), не имеющей этих признаков, при помощи умеренного фага. Так наблюдалась передача свойств разлагать некоторые сахара, образовывать жгутики, споры, устойчивость к пенициллину. Это явление оказалось довольно распространенным среди бактерий. Некоторые созревающие частицы фага захватывают небольшие фрагменты ДНК бактерий, содержащие тот или иной признак, и передают их новой бактерийной клетке - хозяину. Предполагают, что генетический материал донорской клетки частично замещается генетическим материалом умеренного фага во время его пребывания в донорской клетке и этот материал затем фагом переносится в клетку-реципиент. Одновременно трансдуцируются один, реже два тесно сцепленных признака от одной бактериальной клетки к другой.
Конъюгация бактерий. Наличие конъюгации у бактерий наблюдалось некоторыми учеными давно. Но подробно она была изучена в последнее время. Существование конъюгации у бактерий впервые было доказано генетическим методом Ледербергом и Татум (1947). Конъюгация двух бактериальных клеток была обнаружена в электронном микроскопе. Клетки, из которых одна удлиненная - донор, а другая круглая и более крупная - реципиент, сближаются, между ними образуется мостик, по которому генетический материал донора переходит в клетку-реципиент. Был установлен односторонний перенос генетического материала донора, обозначаемого как F + (мужского типа), в клетку-реципиент, обозначаемую знаком F - (женского типа). Клетка-донор содержит особый фактор плодовитости, обозначаемый F-фактор. При скрещивании штаммов F - и F - рекомбинаций не образуется, они образуются при скрещивании штаммов F + и F - с частотой около 10 6 , т. е. очень незначительной.
В дальнейшем были найдены штаммы доноры F + , которые при скрещивании с F - образовывали рекомбинанты с очень большой частотой 10 1 -10 3 . Такие штаммы обозначаются High, frecuency of recombination Hfr. - высокая частота рекомбинации АШ, ЭФ, ЭР. Эти штаммы очень облегчили работу по изучению конъюгации.
При конъюгации происходит медленный перенос генетических факторов бактериальной хромосомы от донорской клетки в реципиентную в строгой последовательности их расположения в хромосоме донора. Количество перенесенных факторов зависит от продолжительности контакта клеток. Конъюгацию можно прекращать искусственно через различные промежутки времени путем встряхивания в особом смесителе, и таким образом можно определять, какие наследственные факторы и в какой последовательности расположены в хромосоме клетки. При помощи конъюгации оказалось возможным легко и более точно составить генетическую карту хромосомы бактерийной клетки.
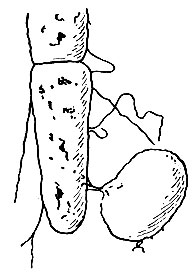
Рис. 48. Электронная микрограмма конъюгации кишечной палочки
Генетика вирусов изучалась главным образом на фагах. Хромосому вируса представляет одна молекула ДНК или РНК, находящаяся в белковой оболочке вируса. Основными признаками фага, наиболее часто используемыми в генетических исследованиях, являются: морфология бляшек на бактерийной культуре, способность фага реагировать с определенными штаммами бактерий хозяев, чувствительность фага к физическим и химическим факторам, лизогения и др. Генетический обмен, скрещивание фагов осуществляется путем заражения одной клетки бактерии двумя генетически различными фагами. В потомстве фага, возникшем в зараженной клетке, обнаруживаются особи с признаками каждого родительского фага и особи с объединенными признаками обоих родителей. Генетический материал фага представляет собой линейно расположенные локусы отдельных признаков. Так были построены генетические карты фагов группы Т и кишечной палочки К-12.
В настоящее время уже встал вопрос о дробимости гена, о его конечных размерах. Так, американец Бензер в опытах по детализации гена r11 фага Т4 разделил этот ген на два цистрона, которые состоят из более мелких единиц: реконов и мутонов. Реконом он считает минимальный участок хромосомы, способный к генетической рекомбинации. Мутон - минимальный участок хромосомы, изменение которого вызывает мутацию. Цистрон - функциональная единица. Он становится неактивным при мутационном повреждении входящего в его состав мутона или рекона. Бензер полагает, что если ген r11 состоит из 3000 нуклеотидов, то мутон его состоит всего из 7 нуклеотидов.
Практическая задача генетики - направленное получение микроорганизмов с полезными для человека свойствами. Значение ее особенно наглядно на примере создания микробов высокопродуктивных культур, образующих антибиотические, физиологически активные вещества, дефицитные аминокислоты и пр. Лучшим методом для этого является селекция мутантов, полученных при воздействии различных мутагенных факторов на исходные (дикие) штаммы. Процесс выведения высокопродуктивных штаммов состоит из многих ступенчатых этапов, состоящих из воздействия на культуру мутагенных факторов с последующим отбором наиболее продуктивного штамма. Этот мутантный штамм может еще подвергаться воздействию мутагенов с последующим отбором наиболее продуктивных мутантов. Так был получен советский штамм пенициллина "Новый гибрид 369", при выведении которого применялись гибридизация, ультрафиолетовые лучи, этиленимин и отбор. Этот штамм применяется на пенициллиновых заводах многих стран.
Во Всесоюзном институте антибиотиков в очень короткий срок была увеличена во много раз активность исходных штаммов при помощи воздействия на них ультрафиолетовых, рентгеновских лучей, химического соединения этиленимина и др. воздействий.
Геном вирусов имеет простое строение и малую молекулярную массу. Число генов у вирусов колеблется от 4—6 (парвовирусы) до 150 генов и больше (вирус оспы). В основе изменчивости вирусов лежат мутации. Мутации носят случайный характер или могут быть направленными. Вирус, являясь облигатным внутриклеточным паразитом, реализует этот паразитизм на генетическом уровне. Присутствие нескольких типов вирусов в инфицированных клетках, т.е. смешанная инфекция, может приводить к таким генетическим взаимодействиям между ними, как множественная реактивация, рекомбинация, кросс-реактивация и др.; могут иметь место и не генетические взаимодействия — комплементация и др.
Множественная реактивация — процесс взаимодействия вирусов с поражением разных генов, в результате которого взаимодействующие вирионы дополняют друг друга благодаря генетической рекомбинации, образуя неповрежденный вирус. Рекомбинация — обмен генетическим материалом между вирусами — возможна в виде обмена генами (межгенная рекомбинация) или участками одного и того же гена (внутригенная рекомбинация). У вирусов рекомбинация происходит в процессе заражения двумя или более типами вирусов, отличающимися друг от друга по генетическим признакам. Вариантом рекомбинации является перекрестная реактивация, или кросс-реактивация, происходящая в том случае, когда у одного из штаммов вируса часть генома повреждена, а другой геном нормальный. При смешанной инфекции двумя такими вирусами в результате рекомбинации появляются штаммы вируса со свойствами родительских микроорганизмов.
В качестве примера негенетического взаимодействия вирусов может быть приведена комплементация: при смешанной инфекции стимулируется репродукция обоих участников взаимодействия или одного из них без изменения генотипов вирусов. Комплементация широко распространена среди вирусов и наблюдается между как родственными, так и неродственными вирусами. Обмен генетическим материалом при этом феномене не наблюдается.
Если геном одного вируса заключен в капсид другого вируса, этот феномен называется фенотипическим смешиванием, наблюдаемым при смешанной инфекции.
Возможны также генетические взаимодействия неродственных вирусов, изучаемые генетической инженерией.
Изучение генетики микроорганизмов не только имеет важное биологическое значение, но и способствует решению многих медицинских проблем, таких, как разработка патогенетических основ лечения и профилактики инфекционных болезней, способов диагностики (полимеразная цепная реакция, ДНК-зонды), создание профилактических, лечебных и диагностических препаратов.
44. Основные группы антимикробных химиопрепаратов, применяемых в терапии и профилактики инфекционных болезней.
В 1932 г. Г. Домагк синтезировал первый сульфаниламидный препарат — стрептоцид, явившийся родоначальником многочисленной группы сульфаниламидных соединений(табл. 8.1), к которым чувствительны ряд грамположительных и грамотрицательных бактерий, прежде всего пиогенные стрептококки, менингококки, гонококки, кишечная палочка и др.
Изучение механизма антибактериального действия сульфаниламидов привело к открытию антиметаболитов— соединений, имеющихьструктурное сходство с важнейшими метаболитами, участвующими в анаболических или катаболических реакциях. Включение антиметаболита в эти реакции приводит соответствующие бактерии к задержке размножения и последующей гибели. Сульфаниламиды оказывают бактериостатическое действие.
Помимо сульфаниламидов, к антиметаболитам относятся аналоги изоникотиновой кислоты, азотистых оснований и других соединений.
Однако антиметаболиты нашли сравнительно ограниченное применение в химиотерапии инфекционных заболеваний.
Это объясняется однотипностью многих биохимических реакций, протекающих в клетках бактерий и человека, поэтому один и тот же антиметаболит блокирует образование продуктов, необходимых для жизнедеятельности микробов и определенных клеток организма человека.
К препаратам, блокирующим процессы репликации и транскрипции относится группа хинолонов.
К препаратам, нарушающим энергетический метаболизм, относятся производные оксихинолина.
Из производных тиосемикарбазона применяется фарингосепт, обладающий бактериостатической активностью в отношении пиогенного стрептококка и других гемолитических стрептококков, встречающихся на миндалинах при ангинах, а также в полости рта при гингивитах и стоматитах.
Презентация на тему: " Генетичекий обмен у бактерий Работа студентки Рябчун Александры." — Транскрипт:
1 Генетичекий обмен у бактерий Работа студентки Рябчун Александры
2 Генетичекий обмен у бактерий процесс передачи генетического материала у бактерий. Основные пути осуществления: -трансформация -трансдукция -конъюгация Конечным этапом генетического обмена, который может быть как внутривидовым, так и межвидовым, является рекомбинация. Рекомбинация процесс взаимодействия между молекулами ДНК,приводящей к формированию новой рекомбинантной молекулы, несущей признаки от бактерии-донора и от бактерии-реципиента.
3 Рекомбинация Законная Требует наличия протяженных комплементарных участков ДНК в рекомбинируемых молекулах Происходит только между близкородственными видами Незаконная Не требует наличия протяженных комплементарных участков ДНК Происходит при участии Is-элементов, обеспечивающих быстрое встраивание в хромосому
5 Трансформация Передача генетического материала между бактериями при помощи фрагментов ДНК. Впервые была воспроизведена Ф.Гриффитсом в 1928 г. Он одновременно вводил в брюшную полость белых мышей авирулентные бескапсульные штаммы пневмококка и убитые капсульные варианты этих бактерий,в результате авирулентные штаммы приобрели вирулентность.
7 Условия, необходимые для успешной трансформации : ДНК донора должна быть выделена из бактериальной культуры того же вида, что и реципиент(или близкородственного) Участок трансформирующей ДНК должен сохранять двунитчатую суперспирализцию Концентрация ДНК не должна быть малой или избыточной, в обоих случаях количество рекомбинантов снижается Клетки-реципиенты должны быть компетентными, т.е. способными адсорбировать на своей поверхности ДНК донора и поглощать ее
8 Стадии трансформации 1.Адсорбция ДНК-донора на клетке- реципиенте 2.Проникновение ДНК внутрь клетки- реципиента 3.Соединение ДНК с гомологичным участком хромосомы реципиента с последующей рекомбинацией
11 Постановка опыта по передаче локуса устойчивости к стрептомицину В опыт берут: ДНК стрептомицинустойчивого штамма Стрептомицинчувствительную культуру в компетентном состоянии Селективную среду, содержащую стрептомицин Последовательность действий: 1. Компетентные клетки реципиента соединяют с ДНК донора и инкубируют в течение 30 минут для контакта. 2.В пробирку вносят раствор фермента ДНК-азы для разрушения не проникшей в реципиентные клетки ДНК. 3.Полученную смесь высевают на чашки с селективной средой и инкубируют. 4.Делают контрольные высевы ДНК донора и культуры- реципиента на селективной среде.
12 Результаты опыта: 1.В обоих контролях рост колоний отсутствует. 2.На опытных чашках вырастают колонии рекомбинантов, которые приобрели признак стрептомицинустойчивости. С помощью данного опыта можно определить частоту трансформации – отношение числа выросших рекомбинантов к числу реципиентных клеток.
13 Трансдукция процесс переноса генетического материала от бактерии-донора к бактерии- реципиенту с помощью бактериофага специфическая неспецифическая (локализованная) (общая) абортивная
14 - бактериофаг переносит любые гены донора; - неспецифическую трансдукцию осуществляют вирулентные фаги; - включение ДНК клетки- рецепиента (фиолетовая) при сборке фага носит случайный характер Неспецифическая трансдукция:
15 Основные этапы: Адгезия на поверхности бактерии-донора с последующим проникновением Размножение бактериофага внутри клетки Самосборка фаговых частиц и образование дефектного бактериофага (сохраняет инфекционные свойства и содержит какой-либо фрагмент ДНК бактерии донора) Перенос дефектным бактериофагом включенной ДНК в клетку-реципиент Рекомбинация и включение перенесенной ДНК в клетку- рециент, а следовательно, изменение ее свойств
16 Специфическая трансдукция фаг переносит определенные гены от бактерии-донора к бактерии-реципиенту Перенос гена lac+ с помощью фага Р1
18 Абортивная трансдукция- трансдукция, при которой перенесенный материал передается только одной из двух дочерних клеток.
19 Основные этапы: Формирование дефектного бактериофага, который содержит фрагменты собственной ДНК и ДНК донора Перенос дефектным бактериофагом включенной ДНК в клетку-реципиент Внесенный фагом фрагмент донорной ДНК не интегрирует в бактериальную хромосому и не реплицируется Однолинейное наследование донорного гена и в конечном итоге утрачивается в потомстве
22 Конъюгация Необходимое условие : наличие в клетке-доноре трансмиссивной плазмиды. Процесс конъюгации у бактерий впервые был обнаружен Джошуа Ледербергом и Эдвардом Тейтумом в 1946 г. форма обмена генетическим материалом между бактериями при их непосредственном клеточном контакте.
25 Основные этапы: Прикрепление клетки-донора к реципиентной клетке с помощью половых ворсинок Образование между обеими клетками конъюгационного мостика Разрыв и деспирализация одной из нитей ДНК, проникновение проксимального конца в клетку- реципиент через конъюгационный мостик Достраивание второй нити ДНК в клетке-реципиенте и восстановление ДНК-донора
26 Типы скрещивания: 1.Скрещивание F + x F - :передается только F-плазмида, при этом F - клетка становится F + -клеткой,приобретая плазмиду и свойства донора. Хромосомные гены не передаются. 2.Скрещивание Hfr x F - : (есть рекомбинанты) передаются бактериальные гены. Для проникновения всей хромосомной нити требуется много времени и, как правило, полный переход осуществляется редко, соответственно, гены, расположенные в той части хромосомы, которая не успела проникнуть в реципиентную клетку, не передаются вообще. Поэтому клетки-реципиенты при таком скрещивании, как правило, не становятся донорами 3.Скрещивание F x F - : (есть рекомбинанты) происходит аналогично скрещиванию F + x F - и реципиентная клетка превращается в донорную
27 Постановка опыта скрещивания Hfr x F - по передаче локусов Pro, Thr, Leu В опыт берут: Донор-штамм с генотипом Pro +, Thr +, Leu +, чувствительный к стрептомицину Реципиент-штамм с генотипом Pro -, Thr -, Leu -, резистентный к стрептомицину Селективную среду, содержащую стрептомицин Последовательность действий: 1.В опытную пробирку вносят культуры донора и реципиента, инкубируют в течение 30 минут 2.Готовят разведения и высевают на селективную среду, инкубируют 3.Делают контрольные высевы культуры донорных и реципиентных клеток на чашки с селективной средой
28 Результаты опыта: 1.На контрольных чашках рост отсутствует 2.На опытной чашке вырастают рекомбинанты С помощью данного опыта можно определить частоту рекомбинаций – отношение числа выросших рекомбинантов к числу участвующих в опыте реципиентных клеток.
| Название | Общая микробиология предмет, задачи, разделы микробиологии, ее связь с другими науками |
| Анкор | Общая микробиология.doc |
| Дата | 28.01.2017 |
| Размер | 409.5 Kb. |
| Формат файла |  |
| Имя файла | Общая микробиология.doc |
| Тип | Документы #180 |
| страница | 8 из 9 |
| Подборка по базе: 18 Общая теория поверхностей второго порядка.doc, Лекция 1. Общая квалиметрия.pdf, Тесты микробиология вирусология иммунология (1).doc, Финансовый менеджмент, задачи, март, 2020.docx, ответы тесты общая гигиена.docx, Быков Цитология и общая гистология.pdf, Экзамен микробиология.doc, Медицинская биология и общая генетика.pdf, Учение о болезни, общая этиология, общий патогенезМетодические м, Вопр экз Общая часть.docx
Рекомбинация (ре + лат. combinatio — соединение) — возникновение новых последовательностей ДНК в результате разрывов и последующих восстановлений ее молекул. В итоге таких изменений ДНК бактерий появляются так называемые рекомбинантные штаммы, или рекомбинанты Наиболее изучены три типа передачи ДНК, отличающиеся друг от друга способом ее транспортировки: трансформация, трансдукция, конъюгация. 39-41. Генетические рекомбинации. Особенности рекомбинативной изменчивости у бактерий и эукариотов. Трансформация и ее стадии. Генетические рекомбинации. Конъюгация, механизмы и этапы конъюгации. F и Hfr – факторы. Генетические рекомбинации. Трансдукция, типы трансдукции. Микроорганизмам, как и клеткам высших организмов свойственны генетические рекомбинации, которые имеют свои особенности. Они определяются прежде всего способом размножения и закономерностями передачи генетического материала. Известно, что генетические рекомбинации у клеток эукариот совершаются в ходе процессов, сопровождающих половое размножение путем реципрокного (взаимного) обмена фрагментами хромосом. При таком обмене генетическим материалом из двух рекомбинирующих родительских хромосом образуются две рекомбинантные хромосомы. Применительно к данным клеткам это означает, что в результате рекомбинаций возникают две рекомбинантные особи. Прокариотам не свойственно половое размножение. Рекомбинация у них происходит в результате внутригеномных перестроек, заключающихся в изменении локализации генов в пределах хромосомы, или при проникновении в клетку реципиента части ДНК донора. Последнее приводит к формированию неполной зиготы — мерозиготы. В результате рекомбинаций в мерозиготе образуется только один рекомбинат, генотип которого представлен в основном генотипом реципиента с включенным в него фрагментом ДНК донора. Вследствие этого реципрокность генетических рекомбинаций у бактерий не может быть выявлена. Рекомбинации подразделяют на законные и незаконные. Законная рекомбинация требует наличия протяженных, комплементарных участков ДНК в рекомбинируемых молекулах. Она происходит только между близкородственными видами микроорганизмов. Генетические рекомбинации происходят при участии ряда ферментов в пределах отдельных генов или групп сцеплений генов. Существуют специальные гес-гены, детермирующие рекомбинационную способность бактерий. Передача генетического материала (хромосомных генов) от одних бактерий к другим происходит путем трансформации, трансдукции и конъюгации, а плазмидных генов — путем трансдукции и конъюгации. Трансформация — непосредственная передача генетического материала (фрагмента ДНК) донора реципиентной клетке. Процесс трансформации бактерий можно подразделить на несколько фаз: 1) адсорбция ДНК-донора на клетке-реципиенте; 2) проникновение ДНК внутрь клетки-реципиента; 3) соединение ДНК с гомологичным участком хромосомы реципиента с последующей рекомбинацией. После проникновения внутрь клетки трансформирующая ДНК деспирализуется. Затем происходит физическое включение любой из двух нитей ДНК донора в геном реципиента. Переносимая ДНК взаимодействует с ДНК реципиента — происходит гомологичная рекомбинация. Трансдукция (от лат. transductio — перенос, перемещение) — передача ДНК от бактерии-донора к бактерии-реципиенту при участии бактериофага. Различают три типа трансдукции: неспецифическую или общую, специфическую и абортивную. Неспецифическая трансдукция. В процессе репродукции фага в момент сборки фаговых частиц в их головку вместе с фаговой ДНК может проникнуть какой-либо фрагмент ДНК бактерии-донора. При этом фаг может утратить часть своего генома и стать дефектным. Принесенный фагом фрагмент ДНК бактерии-донора способен включаться в гомологическую область ДНК клетки-реципиента путем рекомбинации. Таким образом, при неспецифической трансдукции трансдуцирующие фаги являются только переносчиком генетического материала от одних бактерий к другим, поскольку сама фаговая ДНК не участвует в образовании рекомбинантов (трансдуктантов). Специфическая трансдукция характеризуется способностью фага переносить определенные гены от бактерии-донора к бактерии-реципиенту. Это связано с тем, что образование трансдуцирующего фага происходит путем выщепления профага из бактериальной хромосомы вместе с генами, расположенными на хромосоме клетки-донора рядом с профагом. При взаимодействии трансдуцирующих фагов с клетками реципиентного штамма происходит включение гена бактерии-донора вместе с ДНК дефектного фага в хромосому бактерии-реципиента. Бактерии, лизогенированные дефектным фагом, невосприимчивы, как и нее лизогенные клетки, к последующему заражению гомологичным вирулентным фагом. Абортивная трансдукция. При абортивной трансдукции принесенный фагом фрагмент ДНК бактерии-донора не включается в хромосому бактерии-реципиента, а располагается в ее цитоплазме и может в таком виде функционировать. Во время деления бактериальной клетки трансдуцированный фрагмент ДНК-донора может передаваться только одной из двух дочерних клеток, т.е. наследоваться однолинейно и в конечном итоге утрачиваться в потомстве. 42. Плазмиды, их свойства и основные генетические функции. Генетический анализ, принципы составления генетических карт. Генная инженерия. Генетические методы диагностики инфекционных заболеваний. Молекулярная гибридизация, полимеразная цепная реакция. Плазмиды — внехромосомные мобильные генетические структуры бактерий, представляющие собой замкнутые кольца двунитчатой ДНК. По размерам составляют 0,1—5 % ДНК хромосомы. Плазмиды способны автономно копироваться (реплицироваться) и существовать в цитоплазме клетки, поэтому в клетке может быть несколько копий плазмид. Плазмиды могут включаться (интегрировать) в хромосому и реплицироваться вместе с ней. Различают трансмиссивные и нетрансмиссивные плазмиды. Трансмиссивные (конъюгативные) плазмиды могут передаваться из одной бактерии в другую. Среди фенотипических признаков, сообщаемых бактериальной клетке плазмидами, можно выделить следующие: 1) устойчивость к антибиотикам; 2) образование колицинов; 3) продукция факторов патогенности; 4) способность к синтезу антибиотических веществ; 5) расщепление сложных органических веществ; 6) образование ферментов рестрикции и модификации. Некоторые плазмиды находятся под строгим контролем. Это означает, что их репликация сопряжена с репликацией хромосомы так, что в каждой бактериальной клетке присутствует одна или, по крайней мере, несколько копий плазмид. Число копий плазмид, находящихся под слабым контролем, может достигать от 10 до 200 на бактериальную клетку. Для характеристики плазмидных реплико-нов их принято разбивать на группы совместимости. Несовместимость плазмид связана с неспособностью двух плазмид стабильно сохраняться в одной и той же бактериальной клетке. Несовместимость свойственна тем плазмидам, которые обладают высоким сходством репликонов, поддержание которых в клетке регулируется одним и тем же механизмом. Некоторые плазмиды могут обратимо встраиваться в бактериальную хромосому и функционировать в виде единого репликона. Такие плазмиды называются интегративными или эписомами. У бактерий различных видов обнаружены R-плазмиды, несущие гены, ответственные за множественную устойчивость к лекарственным препаратам — антибиотикам, сульфаниламидам и др., F-плазмиды, или половой фактор бактерий, определяющий их способность к конъюгации и образованию половых пилей, Ent-плазмиды, детерминирующие продукцию энтеротоксина. Плазмиды могут определять вирулентность бактерий, например возбудителей чумы, столбняка, способность почвенных бактерий использовать необычные источники углерода, контролировать синтез белковых антибиотикоподобных веществ — бактериоцинов, детерминируемых плазмидами бактериоциногении, и т. д. Существование множества других плазмид у микроорганизмов позволяет полагать, что аналогичные структуры широко распространены у самых разнообразных микроорганизмов. Плазмиды подвержены рекомбинациям, мутациям, могут быть элиминированы (удалены) из бактерий, что, однако, не влияет на их основные свойства. Плазмиды являются удобной моделью для экспериментов по искусственной реконструкции генетического материала, широко используются в генетической инженерии для получения рекомбинантных штаммов. Благодаря быстрому самокопированию и возможности конъюгационной передачи плазмид внутри вида, между видами или даже родами плазмиды играют важную роль в эволюции бактерий. Полимеразная цепная реакция позволяет обнаружить микроб в исследуемом материале (воде, продуктах, материале от больного) по наличию в нем ДНК микроба без выделения последнего в чистую культуру. Для проведения этой реакции из исследуемого материала выделяют ДНК, в которой определяют наличие специфичного для данного микроба гена. Обнаружение гена осуществляют его накоплением. Для этого необходимо иметь праймеры комплементарного З'-концам ДНК. исходного гена. Накопление (амплификация) гена выполняется следующим образом. Выделенную из исследуемого материала ДНК нагревают. При этом ДНК распадается на 2 нити. Добавляют праймеры. Смесь ДНК и праймеров охлаждают. При этом праймеры, при наличии в смеси ДНК искомого гена, связываются с его комплементарными участками. Затем к смеси ДНК и праймера добавляют ДНК-полимеразу и нуклеотиды. Устанавливают температуру, оптимальную для функционирования ДНК-полимеразы. В этих условиях, в случае комплементарное™ ДНК гена и праймера, происходит присоединение нуклеотидов к З'-концам праймеров, в результате чего синтезируются две копии гена. После этого цикл повторяется снова, при этом количество ДНК гена будет увеличиваться каждый раз вдвое. Проводят реакцию в специальных приборах — амплификаторах. ПЦР применяется для диагностики вирусных и бактериальных инфекций. 43. Генетика вирусов. Внутривидовой и межвидовой обмен генетическим материалом. Геном вирусов имеет простое строение и малую молекулярную массу. Число генов у вирусов колеблется от 4—6 (парвовирусы) до 150 генов и больше (вирус оспы). В основе изменчивости вирусов лежат мутации. Мутации носят случайный характер или могут быть направленными. Вирус, являясь облигатным внутриклеточным паразитом, реализует этот паразитизм на генетическом уровне. Присутствие нескольких типов вирусов в инфицированных клетках, т.е. смешанная инфекция, может приводить к таким генетическим взаимодействиям между ними, как множественная реактивация, рекомбинация, кросс-реактивация и др.; могут иметь место и не генетические взаимодействия — комплементация и др. Множественная реактивация — процесс взаимодействия вирусов с поражением разных генов, в результате которого взаимодействующие вирионы дополняют друг друга благодаря генетической рекомбинации, образуя неповрежденный вирус. Рекомбинация — обмен генетическим материалом между вирусами — возможна в виде обмена генами (межгенная рекомбинация) или участками одного и того же гена (внутригенная рекомбинация). У вирусов рекомбинация происходит в процессе заражения двумя или более типами вирусов, отличающимися друг от друга по генетическим признакам. Вариантом рекомбинации является перекрестная реактивация, или кросс-реактивация, происходящая в том случае, когда у одного из штаммов вируса часть генома повреждена, а другой геном нормальный. При смешанной инфекции двумя такими вирусами в результате рекомбинации появляются штаммы вируса со свойствами родительских микроорганизмов. В качестве примера негенетического взаимодействия вирусов может быть приведена комплементация: при смешанной инфекции стимулируется репродукция обоих участников взаимодействия или одного из них без изменения генотипов вирусов. Комплементация широко распространена среди вирусов и наблюдается между как родственными, так и неродственными вирусами. Обмен генетическим материалом при этом феномене не наблюдается. Если геном одного вируса заключен в капсид другого вируса, этот феномен называется фенотипическим смешиванием, наблюдаемым при смешанной инфекции. Возможны также генетические взаимодействия неродственных вирусов, изучаемые генетической инженерией. Изучение генетики микроорганизмов не только имеет важное биологическое значение, но и способствует решению многих медицинских проблем, таких, как разработка патогенетических основ лечения и профилактики инфекционных болезней, способов диагностики (полимеразная цепная реакция, ДНК-зонды), создание профилактических, лечебных и диагностических препаратов. 44. Основные группы антимикробных химиопрепаратов, применяемых в терапии и профилактики инфекционных болезней. В 1932 г. Г. Домагк синтезировал первый сульфаниламидный препарат — стрептоцид, явившийся родоначальником многочисленной группы сульфаниламидных соединений(табл. 8.1), к которым чувствительны ряд грамположительных и грамотрицательных бактерий, прежде всего пиогенные стрептококки, менингококки, гонококки, кишечная палочка и др. Изучение механизма антибактериального действия сульфаниламидов привело к открытию антиметаболитов— соединений, имеющихьструктурное сходство с важнейшими метаболитами, участвующими в анаболических или катаболических реакциях. Включение антиметаболита в эти реакции приводит соответствующие бактерии к задержке размножения и последующей гибели. Сульфаниламиды оказывают бактериостатическое действие. Помимо сульфаниламидов, к антиметаболитам относятся аналоги изоникотиновой кислоты, азотистых оснований и других соединений. Однако антиметаболиты нашли сравнительно ограниченное применение в химиотерапии инфекционных заболеваний. Это объясняется однотипностью многих биохимических реакций, протекающих в клетках бактерий и человека, поэтому один и тот же антиметаболит блокирует образование продуктов, необходимых для жизнедеятельности микробов и определенных клеток организма человека. К препаратам, блокирующим процессы репликации и транскрипции относится группа хинолонов. К препаратам, нарушающим энергетический метаболизм, относятся производные оксихинолина. Из производных тиосемикарбазона применяется фарингосепт, обладающий бактериостатической активностью в отношении пиогенного стрептококка и других гемолитических стрептококков, встречающихся на миндалинах при ангинах, а также в полости рта при гингивитах и стоматитах. 45. Антибиотики. Классификация. Механизмы действия антибактериальных препаратов на микробы. Антибиотики — химиотерапевтические вещества, продуцируемые микроорганизмами, животными клетками, растениями, а также их производные и синтетические продукты, которые обладают избирательной способностью угнетать и задерживать рост микроорганизмов, а также подавлять развитие злокачественных новообразований. За тот период, который прошел со времени открытия П.Эрлиха, было получено более 10 000 различных антибиотиков, поэтому важной проблемой являлась систематизация этих препаратов. В настоящее время существуют различные классификации антибиотиков, однако ни одна из них не является общепринятой. Читайте также:
 Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу. Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу.
Copyright © Иммунитет и инфекции
|