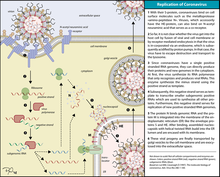
Вирусы репликация которых происходит в ядре
Вирусы — облигатные внутриклеточные паразиты, способные только к внутриклеточному размножению. В вирусинфицированной клетке возможно пребывание вирусов в различных состояниях:
воспроизводство многочисленных новых вирионов;
пребывание нуклеиновой кислоты вируса в интегрированном состоянии с хромосомой клетки (в виде провируса);
существование в цитоплазме клетки в виде кольцевых нуклеиновых кислот, напоминающих плазмиды бактерий.
Поэтому диапазон нарушений, вызываемых вирусом, весьма широк: от выраженной продуктивной инфекции, завершающейся гибелью клетки, до продолжительного взаимодействия вируса с клеткой в виде латентной инфекции или злокачественной трансформации клетки.
Различают три типа взаимодействия вируса с клеткой: продуктивный, абортивный и интегративный.
1. Продуктивный тип — завершается образованием нового поколения вирионов и гибелью (лизисом) зараженных клеток (цитоли-тическая форма). Некоторые вирусы выходят из клеток, не разрушая их (нецитолитическая форма).
Абортивный тип — не завершается образованием новых вирионов, поскольку инфекционный процесс в клетке прерывается на одном из этапов.
Интегративный тип, или вирогения —характеризуется встраиванием (интеграцией) вирусной ДНК в виде провируса в хромосому клетки и их совместным сосуществованием (совместная репликация).
Репродукция вирусов (продуктивный)
1) адсорбция вирионов на клетке;
2) проникновение вируса в клетку;
4) синтез вирусных компонентов;
5) формирование вирионов;
6) выход вирионов из клетки.
У различных вирусов эти стадии отличаются
Специфические рецепторы клеток имеют различную природу, являясь белками, липидами, углеводными компонентами белков, липидов и др. Так, рецепторами для вируса гриппа является сиаловая кислота в составе гли-копротеинов и гликолипидов (ганглиозидов) клеток дыхательных путей. Вирусы бешенства адсорбируются на ацетилхолиновых рецепторах нервной ткани, а вирусы иммунодефицита человека — на СО4-рецепторах Т-хелперов, моноцитов и дендритных клеток. На одной клетке находится от десяти до ста тысяч специфических рецепторов, поэтому на ней могут адсорбироваться десятки и сотни вирионов.
Наличие специфических рецепторов лежит в основе избирательности поражения вирусами определенных клеток, тканей и органов. Это так называемый тропизм (греч. tropos — поворот, направление). Например, вирусы, репродуцирующиеся преимущественно в клетках печени, называются гепатотропными, в нервных клетках — нейротропными, в иммунокомпетентных клетках — иммунотропными и т. д.
Проникновение вирусов в клетку. Вирусы проникают в клетку путем рецептор-зависимого эндоцитоза (виропексиса), или слияния оболочки вируса с клеточной мембраной, или же в результате сочетания этих механизмов.
2. Слияние обточки вириона с клеточной мембраной характерно только для некоторых оболочечных вирусов (парамиксовирусов, ретровиру-сов, герпесвирусов), в составе которых имеются белки слияния. Происходит точечное взаимодействие вирусного белка слияния с липидами клеточной мембраны, в результате чего вирусная липопротеиновая оболочка интегрирует с клеточной мембраной, а внутренний компонент вируса попадает в цитозоль.
В) Синтез вирусных компонентов. Синтез белков и нуклеиновых кислот вируса, который разобщен во времени и пространстве. Синтез осуществляется в разных частях клетки, поэтому такой способ размножения вирусов называется дизъюнктивным (от лат. disjunctus — разобщенный).
С)Синтез вирусных белков. В зараженной клетке вирусный геном кодирует синтез двух групп белков:
1. неструктурных белков, обслуживающих внутриклеточную репродукцию вируса на разных его этапах;
2. структурных белков, которые входят в состав вириона (геномные, связанные с геномом вируса, капсидные и су-перкапсидные белки).
К неструктурным белкам относятся: 1) ферменты синтеза РНК или ДНК (РНК- или ДНК-полимеразы), обеспечивающие транскрипцию и репликацию вирусного генома; 2) белки-регуляторы; 3) предшественники вирусных белков, отличающиеся своей нестабильностью в результате быстрого нарезания на структурные белки; 4) ферменты, модифицирующие вирусные белки, например, протеиназы и протеинкиназы.
I. ДНК-содержашие вирусы реализуют генетическую информацию так же, как и клеточный геном, по схеме:
Причем ДНК-содержашие вирусы используют для этого процесса клеточную полимеразу (вирусы, геномы которых транскрибируются в ядре клетки — аденовирусы, па-повавирусы, герпесвирусы) или собственную РНК-полимеразу (вирусы, геномы которых транскрибируются в цитоплазме, например поксвирусы).
II. Плюс-нитевые РНК-содержашие вирусы (например, пикорнавирусы, флавивирусы, тогавирусы) имеют геном, выполняющий функцию иРНК; он распознается и транслируется рибосомами. Синтез белков у этих вирусов осуществляется без акта транскрипции по схеме:
геномная РНК вируса -> трансляция белка вируса.
III. Геном минус-однонитевых РНК-содержаших вирусов (ортомиксовирусов, парамиксовирусов, рабдовирусов) и двунитевых (реовирусов) служит матрицей, с которой транскрибируется иРНК, при участии РНК-полимеразы, связанной с нуклеиновой кислотой вируса. Синтез белка у них происходит по схеме:
IV. Ретровирусы (вирусы иммунодефицита человека, онкогенные ретровирусы) имеют уникальный путь передачи генетической информации. Геном ретровирусов состоит из двух идентичных молекул РНК, т. е. является диплоидным. В составе ретровирусов есть особый вирусоспецифический фермент — обратная транскриптаза, или ревертаза, с помощью которой осуществляется процесс обратной транскрипции, т. е. на матрице геномной РНК синтезируется комплементарная однонитевая ДНК (кДНК). Комплементарная нить ДНК копируется с образованием двунитевой комплементарной ДНК, которая интегрирует в клеточный геном и в его составе транскрибируется в иРНК с помощью клеточной ДНК-зависимой РНК-полимеразы. Синтез белков для этих вирусов осуществляется по схеме:
Репликация вирусных геномов, т. е. синтез вирусных нуклеиновых кислот, приводит к накоплению в клетке копий исходных вирусных геномов, которые используются при сборке вирионов. Способ репликации генома зависит от типа нуклеиновой кислоты вируса, наличия вирусоспецифических или клеточных полимераз, а также от способности вирусов индуцировать образование полимераз в клетке.
Механизм репликации отличается у вирусов, имеющих:
1) двунитевую ДНК;
2) однонитевую ДНК;
3) плюс-однонитевую РНК;
4) минус-одноните-вую РНК;
5) двунитевую РНК;
6) идентичные плюс-нитевые РНК (ретровирусы).
1. Двунитевые ЛНК-вирусы. Репликация двунитевых вирусных ДНК происходит обычным полуконсервативным механизмом: после рас- плетения нитей ДНК к ним комплементарно достраиваются новые нити. Каждая вновь синтезированная молекула ДНК состоит из одной родительской и одной вновь синтезированной нити. К этим вирусам относится большая группа вирусов, которые содержат двунитевую ДНК в линейной (например, герпесвирусы, аденовирусы и поксвирусы) или в кольцевой форме, как папилломавирусы. У всех вирусов, кроме поксвирусов, транскрипция вирусного генома происходит в ядре.
Уникальный механизм репликации характерен для гепаднавирусов (вируса гепатита В). Геном гепаднавирусов представлен дву-нитевой кольцевой ДНК, одна нить которой короче (неполная плюс-нить) другой нити. Первоначально достраивается (рис. 3.7). Затем полная двунитевая ДНК с помощью клеточной ДНК-зависимой РНК-полимеразы транскрибируется с образованием небольших молекул иРНК и полной однонитевой плюс-РНК. Последняя называется прегеномной РНК; она является матрицей для репликации генома вируса. Синтезированные иРНК участвуют в процессе трансляции белков, в том числе вирусной РНК-зависимой ДНК-полимеразы (обратной транскриптазы). С помощью этого фермента мигрирующая в цитоплазму прегеномная РНК обратно транскрибируется в минус-нить ДНК, которая, в свою очередь, служит матрицей для синтеза плюс-нити ДНК. Этот процесс заканчивается образованием двунитевой ДНК, содержащей неполную плюс-нить ДНК.
Однонитевые ДНК-вирусы. Единственными представителями однонитевых ДНК-вирусов являются парвовирусы. Парвовирусы используют клеточные ДНК-полимеразы для создания двунитевого вирусного генома, так называемой репликативной формы послед него. При этом на исходной вирусной ДНК (плюс-нить) комплементарно синтезируется минус-нить ДНК, служащая матрицей для синтеза плюс-нити ДНК нового вириона. Параллельно синтезируется иРНК, происходит трансляция вирусных пептидов.
Плюс-однонитевые РНК-вирусы. Эти вирусы включают большую группу вирусов — пикорнавирусы, флавивирусы, тогавирусы (рис.3.8), у которых геномная плюс-нить РНК выполняет функцию иРНК. Например, РНК полиовирусов после проникновения в клетку связывается с рибосомами, работая как иРНК, и на ее основе синтезируется большой полипептид, который расщепляется на фрагменты: РНК-зависимую РНК-полимеразу, вирусные протеазы и капсидные белки. Полимераза на основе геномной плюс-нити РНК синтезирует минус-нить РНК; формируется временно двойная РНК, названная промежуточным репликативным звеном. Это промежуточное репликативное звено состоит из полной плюс-нити РНК и многочисленных частично завершенных минус-нитей. Когда образованы все минус-нити, они используются как шаблоны для синтеза новых плюс-нитей РНК. Этот механизм используется как для размножения геномной РНК вируса, так и для синтеза большого количества вирусных белков.
Минус-однонитевые РНК-вирусы. Минус -однонитевые РНК-вирусы (рабдовирусы, парамиксовирусы, ортомиксовирусы) имеют в своем составе РНК-зависимую РНК-полимеразу. Проникшая в клетку геномная минус- нить РНК трансформируется вирусной РНК-зависимой РНК-полимеразой в неполные и полные плюс-нити РНК. Неполные копии выполняют роль иРНК для синтеза вирусных белков. Полные копии являются матрицей (промежуточная стадия) для синтеза минус-нитей геномной РНК потомства
Двунитевые РНК-вирусы. Механизм репликации этих вирусов (реовирусов и ротави-русов) сходен с репликацией минус-однонитевых РНК-вирусов. Отличие состоит в том, что образовавшиеся в процессе транскрипции плюс-нити функционируют не только как иРНК, но и участвуют в репликации: они являются матрицами для синтеза минус-нитей РНК. Последние в комплексе с плюс-нитями РНК образуют геномные двунитевые РНК вирионов. Репликация вирусных нуклеиновых кислот этих вирусов происходит в цитоплазме клеток.
6. Ретровирусы (плюс-нитевые диплоидные РНК-содержащие вирусы). Обратная транс-криптаза ретровирусов синтезирует (на матрице РНК-вируса) минус-нить ДНК, с которой копируется плюс-нить ДНК с образованием двойной нити ДНК, замкнутой в кольцо (рис. 3.10). Далее двойная нить ДНК интегрирует с хромосомой клетки, образуя провирус. Многочисленные вирионные РНК образуются в результате транскрипции одной из нитей интегрированной ДНК при участии клеточной ДНК-зависимой РНК-полимеразы.
Формирование вирусов. Вирионы формируются путем самосборки: составные части вириона транспортируются в места сборки вируса — участки ядра или цитоплазмы клетки. Соединение компонентов вириона обусловлено наличием гидрофобных, ионных, водородных связей и стерического соответствия.
Существуют следующие общие принципы сборки вирусов:
Формирование вирусов — многоступенчатый процесс с образованием промежуточных форм, отличающихся от зрелых вирионов по составу полипептидов.
Сборка просто устроенных вирусов заключается во взаимодействии вирусных нуклеиновых кислот с капсидными белками и в образовании нуклеокапсидов.
У сложноустроенных вирусов сначала формируются нуклеокапсиды, которые взаимодействуют с модифицированными мембранами клеток (будущей липопротеиновой оболочкой вируса).
Причем сборка вирусов, реплицирующихся в ядре клетки, происходит с участием мембраны ядра, а сборка вирусов, репликация которых идет в цитоплазме, осуществляется с участием мембран эндоплазматической сети или плазматической мембраны, куда встраиваются гликопротеины и другие белки оболочки вируса.
У ряда сложноустроенных вирусов минус-нитевых РНК-вирусов (ортомиксовирусов, парамиксовирусов) в сборку вовлекается так называемый матриксный белок (М-белок), который расположен под модифицированной клеточной ембраной. Обладая гидрофобными свойствами, он выполняет роль посредника между нуклеокапсидом и вирусной липопротеиновой оболочкой.
□ Сложноустроенные вирусы в процессе формирования включают в свой состав некоторые компоненты клетки хозяина, например липиды и углеводы.
Выход вирусов из клетки. Полный цикл репродукции вирусов завершается через 5—6 ч (вирус гриппа и др.) или через несколько суток (гепатовирусы, вирус кори и др.). Процесс репродукции вирусов заканчивается выходом их из клетки, который происходит взрывным путем или почкованием, экзоцитозом.
Взрывной путь: из погибающей клетки одновременно выходит большое количество вирионов. По взрывному пути выходят из клетки просто устроенные вирусы, не имеющие липопротеиновой оболочки.
Почкование, экзоцшпт присущи вирусам, имеющим липопротеиновую оболочку, которая является производной от клеточных мембран. Сначала образовавшийся нуклеокапсид или сердцевина вириона транспортируется к клеточным мембранам, в которые уже встроены вирусоспецифические белки. Затем в области контакта нуклеокапсида или сердцевины вириона с клеточной мембраной начинается выпячивание этих участков. Сформировавшаяся почка отделяется от клетки в виде сложно устроенного вируса. При этом клетка способна длительно сохранять жизнеспособность и продуцировать вирусное потомство.
Почкование вирусов, формирующихся в цитоплазме, может происходить либо через плазматическую мембрану (например, парамиксовирусы, тогавирусы), либо через мембраны эндоплазматической сети с последующим их выходом на поверхность клетки (например, буньявирусы).
Вирусы, формирующиеся в ядре клетки (например, герпесвирусы), почкуются в перинуклеарное пространство через модифицированную ядерную мембрану, приобретая таким образом липопротеиновую оболочку. Затем они транспортируются в составе цитоплазма-тических везикул на поверхность клетки.