
Вирус гриппа имеет суперкапсид

По типу строения вирионов выделяют: спиральный тип симметрии (рабдовирусы, вирусы гриппа, парагриппа, коронавирусы); квазисферический – кубический, или икосаэдральный, тип симметрии; cмешанный – у Т-четных бактериофагов (головка в виде многогранника, а хвост в виде спирали).
У вируса табачной мозаики – спиральный тип симметрии вириона. Белковый чехол состоит из отдельных субъединиц в виде шестигранников. Капсомеры – белковые субъединицы, на внутренней поверхности которых расположен желобок, где находится спиралевидная РНК. Капсомеры идентичны друг другу: один и тот же белок, повторяющиеся белковые молекулы (экономится генетический материал). Диаметр капсида 15 – 18 нм, длина вириона до 300 нм (вид палочки). Капсид имеет жесткую структуру.
Тип симметрии определяется только нуклеокапсидом, суперкапсид при этом не учитывается. Например, вирус гриппа снаружи выглядит как сфера, нуклеокапсид имеет спиральный тип симметрии.
Икосаэдр – многогранник, состоящий из 12 вершин и 20 ребер. К данному типу симметрии относятся аденовирусы человека и животных. Капсомеры могут иметь разное строение и содержать разные белки. У аденовирусов боковые капсомеры в виде гексонов, а вершинные – пентонов (соседствуют с пятью капсомерами). От пентамеров отходят выросты (фибры). Это прикрепительные белки. Икосаэдр обеспечивает прочность капсида (защиту от внешних факторов), а прочная связь между капсомерами – минимум свободной энергии.
Многие вирусы имеют суперкапсид – дополнительную оболочку сложно устроенных вирусов, или пеплос. Структурные элементы суперкапсида – шипики, или пепломеры. У вируса гриппа их два типа: гемагглютинины и нейраминидаза; у парагриппа два комплекса – гемагглютинин и нейраминидаза вместе и второй так называемый белок слияния; у вирусов иммунодефицита человека они представлены гликопротеидами. Большинство вирусов, патогенных для человека, являются сложно устроенными. Если у вируса отсутствует суперкапсид, то это просто устроенный вирус.
У большинства сложно устроенных вирусов суперкапсид – это модифицированная (путем встраивания белков вируса) цитоплазматическая мембрана клеток хозяина. Модификация идет путем встраивания шипиков вируса в участки цитоплазматической мембраны (ЦПМ). Исключение составляют поксвирусы, у которых суперкапсид вирусоспецифический, так как имеются собственные гены, ответственные за синтез суперкапсида. Если культивировать один и тот же вирус в разных клетках, получим разные по биохимическому составу суперкапсиды. Шипы выполняют роль прикрепительных белков на поверхности чувствительных клеток. Если их удалить жирорастворителем или детергентом, вирус полностью теряет инфекционную активность.
Биохимия вирусов также хорошо изучена. В состав вириона входят: белки – 70 – 80 %; нуклеиновые кислоты – 4 – 6 % (РНК), 20 – 30 % (ДНК-вирусы); липиды и углеводы в незначительных количествах.
Вирусные белки – полипептиды, которые состоят из обычных левовращающих аминокислот, отличающихся лишь последовательностью построения. Принцип субъединичности построения капсида – повторяющиеся полипептиды небольшой величины – позволяет экономить генетический материал. Если бы капсид был построен из разных белков, требовался бы огромный геном. Так, у вируса табачной мозаики масса капсида 37,2$10 6 дальтон, а для кодирования этого капсида вирусом используется всего 600 нуклеотидов. Весь геном вируса составляет около 6000 нуклеотидов. Некоторые гены вирусов способны кодировать несколько белков со сдвигом рамки. Например, у вируса гриппа 8 генов, а кодируют они 10 вирусных белков.
Капсид за счет упорядоченности капсомеров по сравнению с простой пептидной цепью обладает меньшей свободной энергией. На уровне вирусов действует принцип саморегуляции – самосборки капсида. Принцип самосборки вирусных белков заключается в том, что при определенных условиях (рН, t°) наблюдается спонтанная сборка вирусных белков близких капсидов. При добавлении вирусных нуклеиновых кислот этот процесс упорядочивается.
Вирусные белки подразделяются на структурные и неструктурные. Структурные белки составляют суперкапсид, в составе сердцевины – геномные белки. Неструктурные белки не входят в состав вирусных частиц и обнаруживаются только в зараженной клетке в процессе репродукции. Это ферменты, осуществляющие функции регуляторов, и белки-регуляторы. Аминокислотная последовательность белков вирусов иная, чем у человека, поэтому их можно определять внутри клетки.
Синтез вирусных белков на рибосомах клеток идет по общим законам и регулируется информационной РНК (иРНК), которая образуется на матричных вирусных нуклеиновых кислотах. Белки вирусов выполняют защитную, адресную и регулирующие функции.
Защитная функция – это экранирование нуклеиновой кислоты вируса от химических факторов, нуклеаз и т. д., благодаря чему вирусы существуют тысячи лет.
Адресная функция состоит в проникновении только в нужную чувствительную клетку, а не в любую. Регулирующие функции выполняют внутриклеточные белки вирусов, ферменты, ферментные комплексы.
Вирусные нуклеиновые кислоты имеют существенные отличия от нуклеиновых кислот всех других существ. Обычно генетическая информация закодирована в двуспиральной ДНК и имеется однонитевая РНК (информационная, транспортная, рибосомальная). У вирусов в качестве геномной может быть как ДНК, так и РНК. У некоторых вирусов (РНК-геномных) вирионная РНК одновременно может выполнять роль информационной. Такие вирусы называют нитевыми. Если выделить в чистом виде такую РНК и поместить в клетку, то инфекция будет протекать так же, как если бы туда проник целый вирус. У других вирусов (нитевых) РНК не может выполнять функцию информационной.
Вирусы отличает многообразная структура нуклеиновых кислот. Их можно получить, разрушив вирус химическими (фенолом) или физическими (ультразвуком) факторами. Выделяют ДНК-содержащие вирусы, у которых она может присутствовать в виде: классической двунитевой (аденовирусы, герпесвирусы); двунитевой линейной с замкнутыми концами (оспа); двунитевой линейной с разрывами одной цепи (Т-фаги); с несколькими разрывами одной нити (каждый фрагмент – уникальный ген); двунитевой, замкнутой в кольцо со сверхвитками, (суперспирализация) или без них (тогавирусы); двунитевой, у которой внешняя нить замкнута в кольцо (L-нить), а у внутренней 1 /3 отсутствует (S – нить, шорт) – гепаднавирусы; уникальной линейной однонитевой ДНК (парвовирусы); однонитевой замкнутой в кольцо (фаги).
РНК-содержащие вирусы также делятся на несколько типов: классический однонитевой линейный (пикорновирусы, тогавирусы, парамиксовирусы, рабдовирусы); линейный однонитевой фрагментированный (ортомиксовирусы); однонитевой фрагментированный, где каждый фрагмент замкнут в кольцо (буньявирусы); двунитевой с идентичными нитями (ретровирусы); двунитевой фрагментированный (реовирусы).
По химическому составу вирусные РНК и ДНК аналогичны клеточным, но в отличие от них содержат метилированный урацил.
Липиды имеют сложно устроенные вирусы, их наличие характерно для патогенных вирусов. Содержание липидов в вирусах различно: от 1,5 до 54 % (тогавирусы). Липидный состав вирусов непостоянен, у большинства это липиды клеточного происхождения. Липиды содержатся в суперкапсиде и зависят от клетки хозяина. Липидный состав различных чувствительных клеток при включении в них одного и того же вируса будет разным, тогда как одинаковые клетки при включении в них разных вирусов сохранят идентичность. Липиды выполняют важную защитную функцию, укрепляя белковый скелет суперкапсида. Они имеют вид липопротеидного или гликопротеидного комплекса. У поксвирусов синтезируются собственные липиды под контролем самого вируса, и поэтому состав липидов у них постоянен.
Углеводы входят в суперкапсид сложно устроенных вирусов преимущественно в составе различных шипов в виде моносахаридов, аминосахаридов (2 – 3 цепочки в комплексе с белковым или липидным компонентом – гликопротеидом, гликолипопротеидом). У вируса гриппа это гемагглютинин и нейраминидаза, у парагриппа – комбинированный шип гемагглютинина и нейраминидазы, у ВИЧ – поверхностные структуры gp 41- и gp 120-гликопротеиды, у вируса клещевого энцефалита – шипы гемагглютинина.
Углеводный компонент вируса определяется клеткой хозяина, укрепляет суперкапсидную структуру, придавая ей жесткость. Удаление гликопротеидных комплексов у сложно устроенных вирусов ведет к потере способности к адсорбции на чувствительных клетках. У просто устроенных вирусов углеводный компонент отсутствует.
В составе вирусов могут быть и другие компоненты. В состав полипептидов, например, часто включаются фосфаты (аденовирусы, ретровирусы, герпесвирусы, поксвирусы, ортомиксовирусы), их конкретная функция пока точно не установлена. Некоторые вирусы содержат микроэлементы: медь, молибден, а отдельные вирусы – целый набор ферментов: поксвирусы – 10, ВИЧ – 4, вирус гриппа – 3 фермента. Ферменты обеспечивают вирусную репродукцию: реакцию полимеризации (образования иРНК), репликацию (образование новых нитей нуклеиновых кислот) по принципу комплементарности. В ряде случаев ферменты синтезируются за счет генома вируса.
Вирусы также активно используют клеточные ферменты, например, у гриппа гемагглютинин находится в составе шипов в неактивном состоянии и для адсорбции на клеточном эпителии необходима протеолитическая активация гемагглютинина за счет ферментов клеточного секрета. Только после этого достигается соответствие прикрепительного белка структурам чувствительной клетки.
С поверхностными белками (H, N) связана высокая антигенная изменчивость вирусов гриппа. Она определяет рецидивы эпидемической агрессивности вируса типа А, меньше проявляется у вируса типа В и, по-видимому, не имеет практического значения для гриппа С (см. ниже).
Вирус проникает в клетку путем эндоцитоза после связывания гемагглютинина с сиалированными клеточными рецепторами. Фагосома (эндосома), в которую заключен вирус, сливается с лизосомами. Кислая среда внутри фаголизосомы меняет конформацию гемагглютинина, обнажая пептиды, вызывающие слияние вирусной и эндосомальной мембран. Благодаря этому нуклеокапсид освобождается от суперкапсида и выходит в цитоплазму. Отсюда вместе с М-белком он быстро транспортируется в ядро, где уже через несколько минут после заражения появляются первые РНК-транскрипты. Число разновидностей мРНК соответствует количеству сегментов геномной РНК, хотя спектр синтезируемых белков несколько больше: к структурным белкам вириона добавляется 3 неструктурных белка, принимающих участие в репликации вируса. Расширение генетической емкости РНК достигается благодаря изящному механизму — повторной трансляции одной и той же мРНК со сдвигом рамки считывания ее генетического кода.
Ортомиксовирусы едва ли не единственные из РНК-вирусов, манипулирующие со своей геномной молекулой в ядре зараженной клетки. Клетки не умеют транскрибировать РНК, поэтому операции такого рода обязан производить сам вирус. Толчок дает полимеразный комплекс вириона, позже начинают работать вновь синтезированные ферменты. Белки РВ1 и РВ2 обеспечивают копирование геномных фрагментов с образованием неполных и полных по длине транскриптов. Укороченные копии выполняют функции мРНК. Для этого они подвергаются доработке, обретая облик, привычный для мРНК клетки (присоединение к 5`-концу шапочки из метилированных нуклеотидов и полиаденилирование 3`-конца). Не располагая собственными ресурсами для прямого решения этой задачи, вирус находит обходной путь. Он подключает свою эндонуклеазу (возможно, белок РА), которая откусывает недостающие фрагменты от мРНК клетки-хозяина и переносит их на вирусные РНК-транскрипты. Столь необычное и тонкое проявление молекулярного паразитизма объясняет природу внутриядерной репликации вирусов гриппа: только здесь они могут найти свежие (еще не связавшиеся с белком) мРНК, чувствительные к вирусной эндонуклеазе.
Полные по длине РНК-транскрипты не модифицируются и поэтому не могут работать как мРНК. Они остаются в ядре и служат матрицей для образования репликативных РНК, идущих на построение новых нуклеокапсидов. В этом принимает участие РНК-полимераза, образующаяся при помощи первых мРНК.
Готовые мРНК переправляются в цитоплазму, где транслируются на рибосомах в вирусные белки. Часть синтезированных белков вновь переходят в ядро и после комплексации с репликативными РНК образуют нуклеокапсиды. Гемагглютинин и нейраминидаза включаются в плазматическую мембрану, готовя фрагменты будущего суперкапсида. С внутренней стороны липидного бислоя к ним присоединяется М-белок, который ориентирует закрепление нуклеокапсидов в зонах почкования новых вирионов. При отделении от клеток вирус получает суперкапсид с его важнейшими атрибутами — гемагглютинином и нейраминидазой (рис. 4, 5).
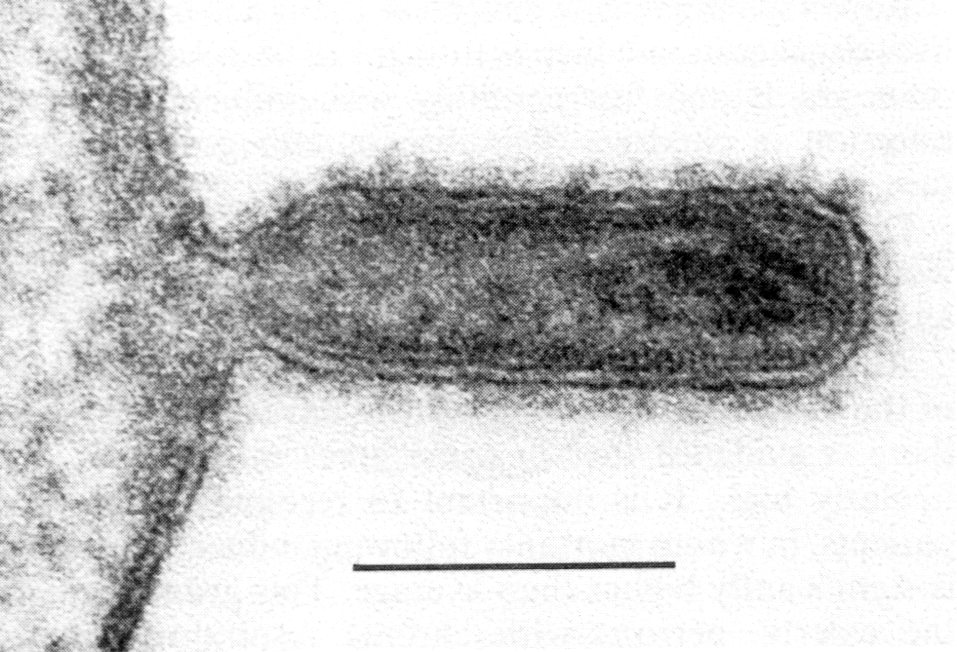
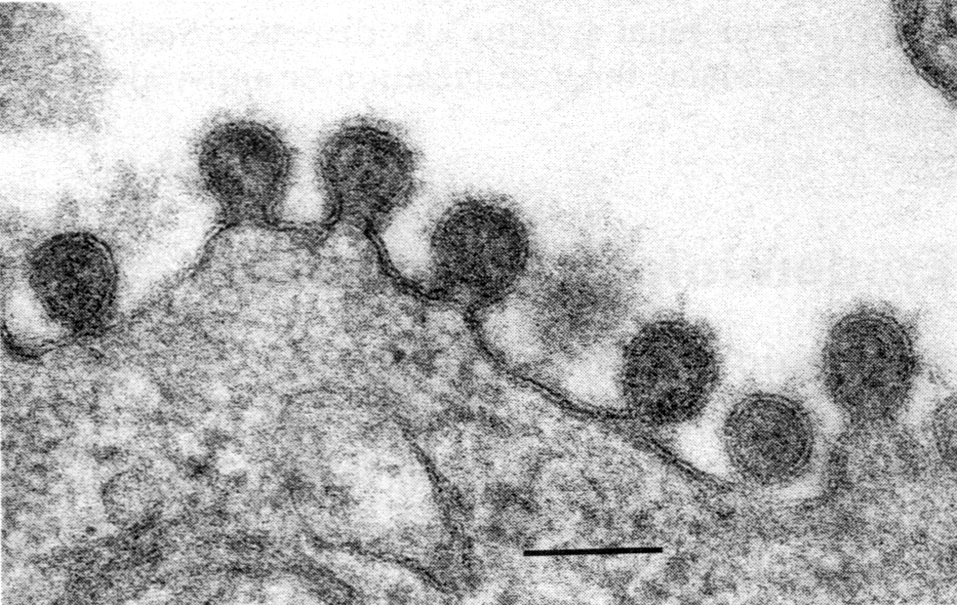
Рис. 4. Почкование вируса гриппа (палочковидные и сферические частицы). Электронограмма. Полоска
соответствует 100 нм 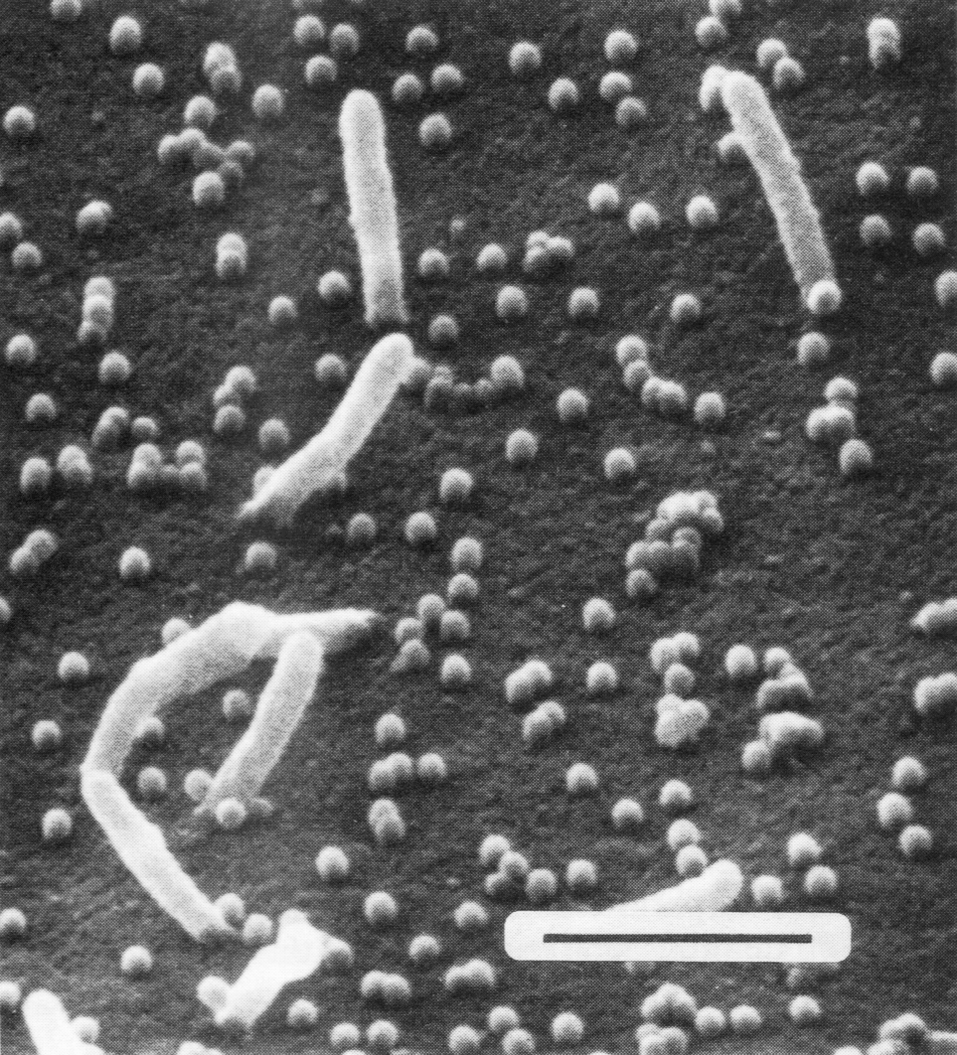
Рис. 5. Сканирующая электронограмма почкующихся вирусов гриппа. Удлиненные структуры являются микроворсинками клетки, почкующиеся вирионы выглядят как сферические частицы. Полоска соответствует 1 мкм
Как уже говорилось, гемагглютинин синтезируется в виде предшественника, который активируется клеточными протеазами (здесь корень формальной логики, породившей иллюзию о возможности лечения вирусных инфекций банальными ингибиторами протеаз). Сбрасывание созревших вирионов и их распространение на соседние клетки усиливается нейраминидазой. Репликация в чувствительных клетках происходит очень быстро, и уже через 6—8 ч в среде появляется вирусное потомство.
Впрочем, в последние годы зарегистрированы случаи заражения людей птичьими штаммами вируса А. Реальная тревога возникла в 1997 г. в Гонконге, когда 18 человек напрямую заразились от домашних птиц вирусом H5N1; 6 человек погибло. В 2003 г. высоковирулентные штаммы птиц H5N1 и H7N7 спровоцировали фатальные случаи гриппа среди жителей Южной Азии. К счастью, они не получили распространения, но тем не менее очертили круг задач, решение которых необходимо для профилактики новых гриппозных пандемий.
Вирусы типов В и С являются сугубо человеческими и в естественных условиях не инфицируют других хозяев.
Вирусы гриппа проникают в респираторный тракт с каплями аэрозоля и частицами пыли. Чем мельче их величина, тем глубже проникает вирус; самые мелкие частицы достигают бронхиол и альвеол. Главным местом размножения вируса служат реснитчатые и бокаловидные клетки слизистой оболочки верхних дыхательных путей, но инфекция может охватить весь респираторный тракт — от носовых ходов до терминальных бронхиол и альвеол. Благодаря короткому репликативному циклу из каждой вирусной частицы к концу первых суток возникает огромное потомство — 10 27 . Это объясняет короткий инкубационный период (1—3 сут) и острейшее начало гриппа.
Повреждение зараженных эпителиоцитов индуцирует воспалительную реакцию, в которой преобладают макрофаги и лимфоциты. Подвергаясь активации, они секретируют множество флогогенных начал (прежде всего цитокинов), которые вызывают общую интоксикацию и поддерживают местный воспалительный процесс. В типичных случаях после внезапного недомогания, озноба с быстрым повышением температуры до 38—40 о С, головной и часто мышечных болей развиваются симптомы поражения верхних (ринит, фарингит) и глубжележащих (ларингит, трахеобронхит) дыхательных путей. Общая интоксикация характерна для гриппа, но часто доминируют местные проявления. Наиболее тяжело протекает грипп А (хотя история эпидемий убеждает в вариабельности этого признака); вирусы типа В и, особенно, С менее агрессивны. В целом, опираясь только на клинику, отличить грипп от других ОРЗ практически невозможно. Без лабораторного подтверждения это удается лишь в сочетании с эпидемиологическим диагнозом, на фоне вспышки гриппа среди населения.
Читайте также:
