Транскрипция вирусной днк это
Известны три вида процессов, в рамках которых осуществляется специализированный перенос генетической информации. Один из них - перенос информации от РНК к РНК - удается зафиксировать только в клетках, зараженных вирусами, генетический материал которых представлен РНК. Это, в частности, вирус табачной мозаики и многие другие вирусы растений, РНК-содержащие бактериофаги и некоторые другие вирусы животных, такие как полиовирусы. Эти вирусные геномные РНК, одноцепочечные или двухцепочечные, несут гены, кодирующие специфические РНК-репликазы, которые по РНК-матрице могут синтезировать комплементарные молекулы РНК, способные в свою очередь служить матрицами для синтеза аналогичным способом копий родительских цепей РНК. Перенос генетической информации от РНК к РНК также основан на принципе комплементарности оснований в родительской и дочерней цепях РНК.
Обратная транскрипция.
Данный вид специализированного переноса генетической информации от РНК к ДНК обнаружен в клетках животных, инфицированных вирусами определенного типа. Это особый тип РНК- содержащих вирусов, называемых ретровирусами. В настоящее время установлено, что еще один тип вирусов - ДНК-содержащий вирус гепатита В - в своем развитии также использует перенос информации от РНК к ДНК.
Ретровирусы содержат молекулы одноцепочечной РНК, при этом каждая вирусная частица имеет две копии РНК-генома, т. е. вирусы этого типа являются единственной известной разновидностью диплоидных вирусов. Впервые они были обнаружены по способности вызывать образование опухолей у животных. Первый вирус этого типа был описан в 1911 г. Пейтоном Раусом, обнаружившим инфекционную саркому у кур.
После проникновения РНК ретровируса в клетку хозяина вирусный геном подвергается обратной транскрипции. При этом сначала образуется дуплекс РНК-ДНК, а затем двухцепочечная ДНК. Эти этапы предшествуют экспрессии вирусных генов на уровне белков и образованию РНК-геномов.
Фермент, катализирующий комплементарное копирование РНК с образованием ДНК, называется обратной транскриптазой. Он содержится в ретровирусных частицах (вирионах) и активизируется после попадания вируса в клетку и разрушения его липидно-гликопротеиновой оболочки.
Имеются данные о том, что обратная транскрипция происходит и в самых разных эукариотических клетках, а обратная транскриптаза играет важную роль в процессах перестройки генома.
Обратные транскриптазы ретровирусов -- это по существу ДНК- полимеразы, которые могут использовать in vitro в качестве матрицы ДНК.
Однако гораздо эффективнее они работают на РНК. Как и все ДНК- полимеразы, обратные транскриптазы не способны инициировать синтез новых цепей ДНК. Но если синтез уже инициирован с помощью праймер- ной РНК или З'-концевого участка ДНК, то фермент эффективно осуществляет синтез, используя цепь ДНК как матрицу.
Ретровирусы оказались очень полезным инструментом современных генно-инженерных исследований. Они служат источником для получения практически чистой обратной транскриптазы - фермента, играющего важнейшую роль в многочисленных работах, основанных на клонировании эукариотических генов. Так, очищенную индивидуальную мРНК, кодирующую интересующий исследователя белок, как правило выделить гораздо легче, чем фрагмент ДНК генома, кодирующий этот белок. Затем с помощью обратной транскриптазы можно получить ДНК-копию этой мРНК и встроить ее в подходящую плазмиду для клонирования и выработки значительных количеств нужной ДНК.
Трансляция ДНК. Третий вид специализированного переноса генетической информации от ДНК непосредственно к белку удалось наблюдать только в лаборатории in vitro. В лабораторных условиях некоторые антибиотики, в частности стрептомицин и неомицин, взаимодействующие с рибосомами, могут так изменять их свойства, что рибосомы начинают использовать в качестве матрицы вместо мРНК одноцепочечную ДНК, с которой последовательность оснований непосредственно переводится в аминокислотную последовательность синтезируемого полипептида.
ДНК-СОДЕРЖАЩИЕ ВИРУСЫ И ФАГИ
Вирусы — это внеклеточная форма жизни, обладающая собственным геномом и способная к воспроизведению только в клетках живых организмов.
Вирион (или вирусная частица) состоит из одной или нескольких молекул ДНК или РНК, заключенных в белковую оболочку (капсид), иногда содержащую также липидные и углеводные компоненты
Говорят: в. это переход от химии к живому
Вирусы являются внутриклеточными паразитами на генетическом уровне и используют для своего размножения белок-синтезирующий аппарат клетки-хозяина.
Жизненный цикл вируса начинается
с проникновения внутрь клетки.
Для этого он связывается со специфическими рецепторами на ее поверхности и
а) либо вводит свою нуклеиновую кислоту внутрь клетки, оставляя белки вириона на ее поверхности,
3. В результате этой процедуры вирусный геном становится доступным для ферментных систем клетки, обеспечивающих экспрессию генов вируса.
4. Именно после проникновения вирусной геномной нуклеиновой кислоты в клетку заключенная в ней генетическая информация расшифровывается генетическими системами хозяина и используется для синтеза компонентов вирусных частиц.
По сравнению с геномами других организмов вирусный геном относительно мал и кодирует лишь ограниченное число белков, в основном белки капсида и один или несколько белков, участвующих в репликации и экспрессии вирусного генома. Необходимые метаболиты и энергия поставляются хозяйской клеткой.
ДНК-содержащие вирусы несут в качестве генетического материала либо одно-, либо двухцепочечную ДНК, которая может быть как линейной, так и кольцевой. В ДНК закодирована информация о всех белках вируса. Вирусы классифицируют в зависимости от того, одно или двухцепочечная у них ДНК, и про- или эукариотической является клетка-хозяин. Вирусы заражающие бактерии называются бактериофагами.
1 — вирусы оспы; 2 — вирусы герпеса; 3 – аденовирусы; 4 — паповавирусы; 5 — гепаднавирусы; 6 — парвовирусы;
Первая группа — вирусы с двуцепочечной ДНК,
- они получили название ретроидные вирусы.
- п редставителями этой группы вирусов являются вирус гепатита В и вирус мозаики цветной капусты.
1. Репликация ДНК-генома этих вирусов осуществляется при посредстве промежуточных молекул РНК:
2. Молекулы РНК образуются в результате транскрипции вирусных ДНК в клеточном ядре хозяйским ферментом ДНК-зависимой РНК-полимеразой.
3. Транскрибируется только одна из нитей вирусной ДНК.
4. Синтез ДНК на РНК-матрице происходит в результате реакции, катализируемой обратной транскриптазой; сначала синтезируется (-) нить ДНК,
5. а затем на вновь синтезированной (-) нити ДНК тот же фермент строит (+) нить.
В целом общая схема репликации генома ретроидных вирусов поразительно похожа на схему репликации генома ретровирусов. По-видимому, данное сходство имеет под собой и эволюционную основу, так как первичная структура обратных транскриптаз этих вирусов выявляет определенное сходство между собой.
Вторая группа — вирусы с двуцепочечной ДНК,
- репликация осуществляется по схеме ДНК -> ДНК.
- с генома этих вирусов в зараженной клетке ДНК-зависимая РНК-полимераза транскрибирует молекулы мРНК (т.е. (+) РНК),
- мРНК (т.е. (+) РНК) принимает участие в синтезе вирусных белков,
- размножение вирусного генома осуществляет фермент ДНК-зависимая ДНК-полимераза: (±) днк → (+)РНК
В одних случаях производством как мРНК, так и ДНК занимаются клеточные ферменты; в других случаях вирусы используют собственные ферменты. Бывает, что те и другие ферменты обслуживают процесс репликации и транскрипции. К этой группе относятся вирусы герпеса, оспы и др.
Оспа – естественный враг СПИДа (нет оспы – есть СПИД). О СПИДе есть информация в Ветхом завете. В нашем геноме есть генетические метки прежних пандемий СПИДа
Вирусные заболевания периодичны: ОСПА → ПРОКАЗА→ЧУМА → †
Третья группа — вирусы с одноцепочечной ДНК, либо с негативной, либо с позитивной полярностью.
- Попав в клетку, вирусный геном сначала превращается в двуцепочечную форму,
- это превращение обеспечивает клеточная ДНК-зависимая ДНК-полимераза:
Транскрипция и репликация на последующих этапах происходит так же, как и для вирусов, с (±) ДНК-геномом.
- обычно встречаются у нитевидных вирусов.
- образуются путем самосборки асимметричных белковых субъединиц (капсомеров), объединяющихся в трубчатую структуру со спиральной симметрией (например у Pf1).
- Субъединицы в большинстве случаев гомогенны, так, что поверхность вириона состоит из множества копий одного и того же белка, хотя под наружним капсидом могут находиться и другие белки.
- ДНК в таких вирусах либо вытянута, либо может быть туго скручена в комплексе со специальными связывающими белками.
- свойственны большинству сферических ДНК-содержащих вирусов
- икосаэдр – это многогранник с двадцатью треугольными гранями, имеющий кубическую симметрию и приблизительно сферическую форму.
- Вершины треугольников соединяясь образуют двенадцать вершин икосаэдра;
- в местах соединения располагаются обычно пентамерные белковые структуры – пентоны; там же могут находится участки, на которых формируются белковые нити, нередко ассоциированные с вершинами ( например у ф Х174 прозрачка 1).
- Грани икосаэдра заполнены другими белковыми субъединицами, сгруппированными обычно в гексамерные структуры – гексоны (апример, у аденовируса прозрачка 1).
- Количество субъединиц, необходиимое для заполнения граней, определяется размерами вириона в целом, и разные икосаэдрические вирусы содержат поэтому разное число гексонов – обычно при неизменном числе пентонов.
- ДНК обычно плотно свернута внутри капсида;
- иногда она связана с белками или полипептидами, способными стабилизировать ее структуру.
Сложные капсиды без оболочки
- типичны для бактериофагов:
- они состоят из частей с разными типами симметрии.
Сложные капсиды с оболочкой
- есть только у вирусов эукариотических клеток.
- В них ДНК-белковые комплексы окружены одним или несколькими белковыми слоями и наружней мембраной, почти все белковые компоненты которой являются вирусными по своему происхождению, а липидные структуры – клеточными.
- зараженные вирусом клетки либо остаются живыми (тогда говорят, что вирус невирулентен),
- либо подвергаются лизису, приводящему к высвобождению вирусных частиц.
- Неизменным итогом заражения клеток ДНК-содержащими бактериофагами является лизис.
- ДНК-содержащие вирусы животных вызывают лизис редко; однако клетки могут погибнуть из-за возникших при заражении хромосомных повреждений, вследствии иммунологической реакцииорганизма или просто в результате нарушения вирусом нормальных клеточных функций.
Размножение вируса – четко очерченный цикл, приводящий в конечном счете, после синтеза новых молекул вирусных белков и большого числа копий вирусной ДНК, к формированию зрелых вирусных частиц. У вирусов бактерий весь цикл может завершаться менее чем за час, тогда как у многих вирусов животных он занимает не один день.
Адсорбция вируса на клетке-хозяине – первый этап инфицирования. Она происходит на специфических рецепторных участках (белковых или липидных) клеточной поверхности, которые узнаются особыми выступающими частями вириона и к которым он прочно прикрепляется. У вирусов без оболочки такими частями могут быть белковые отростки (например, у аденовируса и бактериофага Т2), а у вирусов с оболочкой это, как правило белки, погруженные в вирусную мембрану. В процессе адсорбции осуществляются, в частности, такие белок-белковые взаимодействия, результатом которых является инициация стадии проникновения ДНК в клетку.
Транскрипция и репликация генетического материала вируса осуществляется обычно с участием ферментов клетки-хозяина. Сначала вирусная ДНК копируется РНК-полимеразами клетки-хозяина, в результате чего образуется мРНК, которая затем транслируется. На некоторых молекулах вирусной ДНК синтезируются также ее ДНК-копии – с помощью либо клеточной, либо кодируемой вирусом ДНК-полимеразы. Эти ДНК-копии используются в последствии при сборке вирусных частиц. В некоторых случаях, например, у бактериофага Т4 первые же новосинтезированнные молекулы вирусной мРНК транслируются с образованием специальных белков, модифицирующих полимеразы клетки-хозяина таким образом, что те прекращают транскрипцию клеточных генов, не теряя пари этом способности транскрибировать вирусные. В какой части клетки протекают процессы транскрипции и репликации вирусной ДНК эукариотических вирусов: в ядре или цитоплазме? У одних транскрипция и репликация происходит в ядре клетки-хозяина ( например, у вируса герпеса), а у других – в цитоплазме, например, у поксивирусов.
Трансляция вирусной мРНК на рибосомах клетки-хозяина приводит к образованию вирусных белков. Некоторые из этих белков используются в последствии для построения капсидов, другие связываются с вирусной ДНК, стабилизируя ее (у многих вирусов животных), третьи хотя и не войдут никогда в состав зрелых вирионов, участвуют в процессе их сборки в качестве ферментов (например, у бактериофага Т2)
Сборка вируса из его компонентов в клетке-хозяине может происходить спонтанно (самосборка), но может зависеть и от участия вспомогательных белков. Вирусная ДНК обычно покрывается слоем белка – капсидом. Капсид, в свою очередь, может заключаться в мембранную структуру, получаемую вирионом обычно от клетки-хозяина: покидая клетку путем отпочковывания от нее, вирусная частица оказывается окруженной плазматической мембраной.
ДНК-содержащие опухолеродные вирусы разделяются на следующие 5 классов.
1. Репликация (от ДНК к ДНК)
2. Транскрипция (от ДНК к РНК)(
3. Трансляция (от РНК к белку)
4. Обратная транскрипция (от РНК к кДНК)
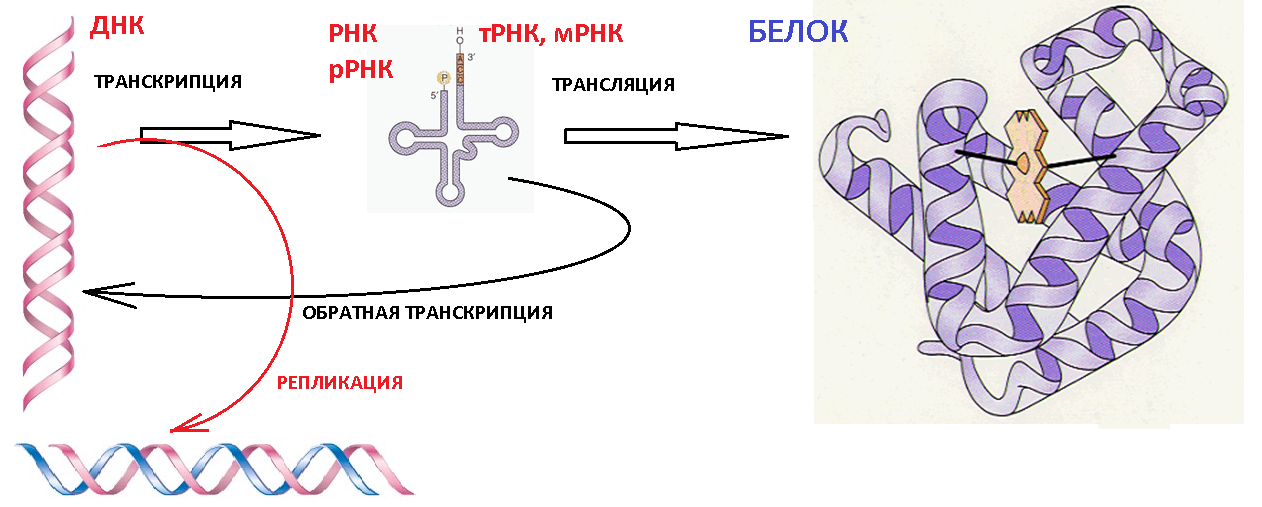
Долгое время считалось, что передача информации от РНК к ДНК невозможна, однако, впоследствии выяснилось, что это не так. Некоторые вирусы способны встраивать информацию со своей вирусной РНК в ДНК генома клетки-хозяина. Возможность "обратного" направления информации в настоящее время все шире используется в различных целях, от исследовательских до терапевтических. Так называемые энзимы- реверс-транскриптазы- способны осуществлять синтез кДНК на матрице РНК. О происходящих в клетках млекопитающих (эукариот) процессах передачи информации известно достаточно много, но далеко не все, и изложение хотя бы известных на данный момент времени сведений потребовалось бы слишком много места. Поэтому далее будут изложены лишь самые основы протекающих в клетках простейших организмов (прокариот) этапов передачи наследственной информации.
В процессе копирования информации происходит синтез дочерних молекул ДНК на основе информации, "записанной" в родительской молекуле ДНК. Ясно, что дочерние молекулы должны представлять собой точные копии родительской.
Репликация может осуществляться тремя способами:
а) консервативным;
б) полуконсервативным;
в) дисперсивным.
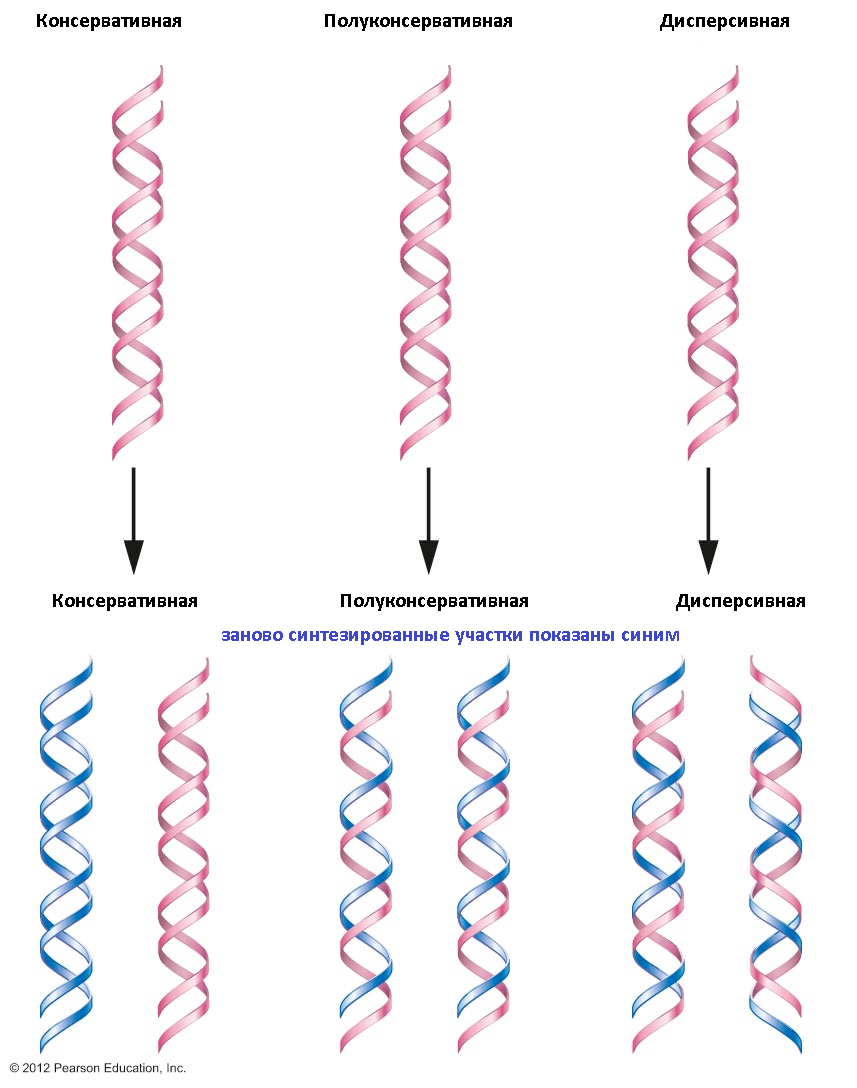
При консервативной репликации вновь синтезированные цепи ДНК находятся в дочерней молекуле. При полукон сервативной репликации полученные молекулы состоят из родительской и вновь синтезированной цепей. Дисперсивный способ репликации означает наличие перемежающихся родительских и вновь синтезированных участков на каждой из цепей образованных молекул ДНК. Для животных организмов и человека ХАРАКТЕРЕН только ПОЛУКОНСЕРВАТИВНЫЙ путь РЕПЛИКАЦИИ ДНК.
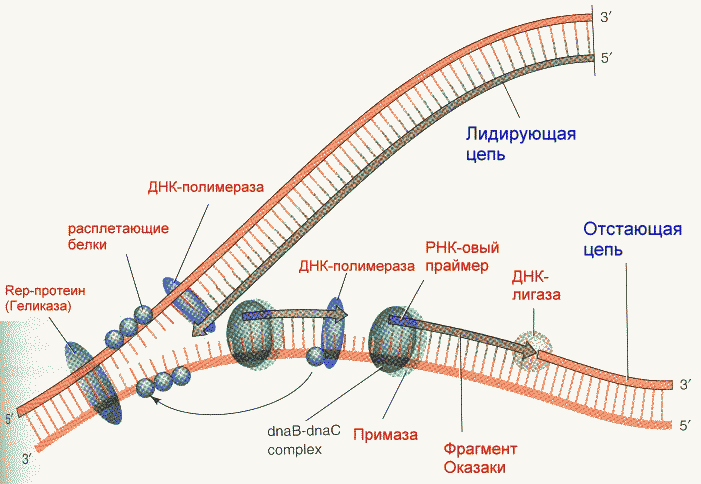
В процессе репликации участвует целый ряд энзимов (ферментов) с определенными функциями. Только синтезирующих ферментов в клетках прокариот насчитывается три. Их называют ДНК- полимеразами I, II и III. Сведения о функциональных особенностях ДНК-полимераз приведены в таблице.
| Проявляемая функция (активность) | |
| Полимеразная 5'-3' | |
| Экзонуклеазная 3'-5' | |
| Экзонуклеазная 5'-3' | |
| молекул в клетке | |
| производительность (нуклеотидов в мин, 37 С, на 1 молекулу Pol) |
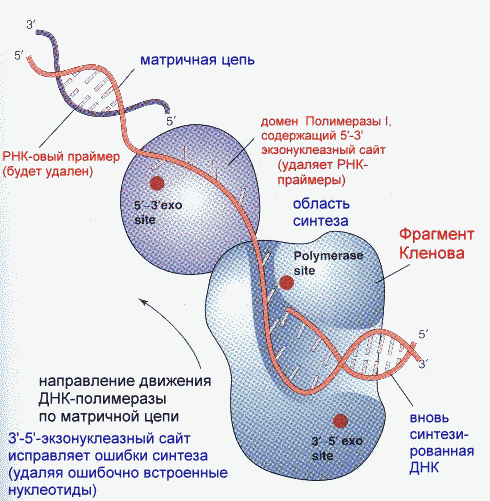
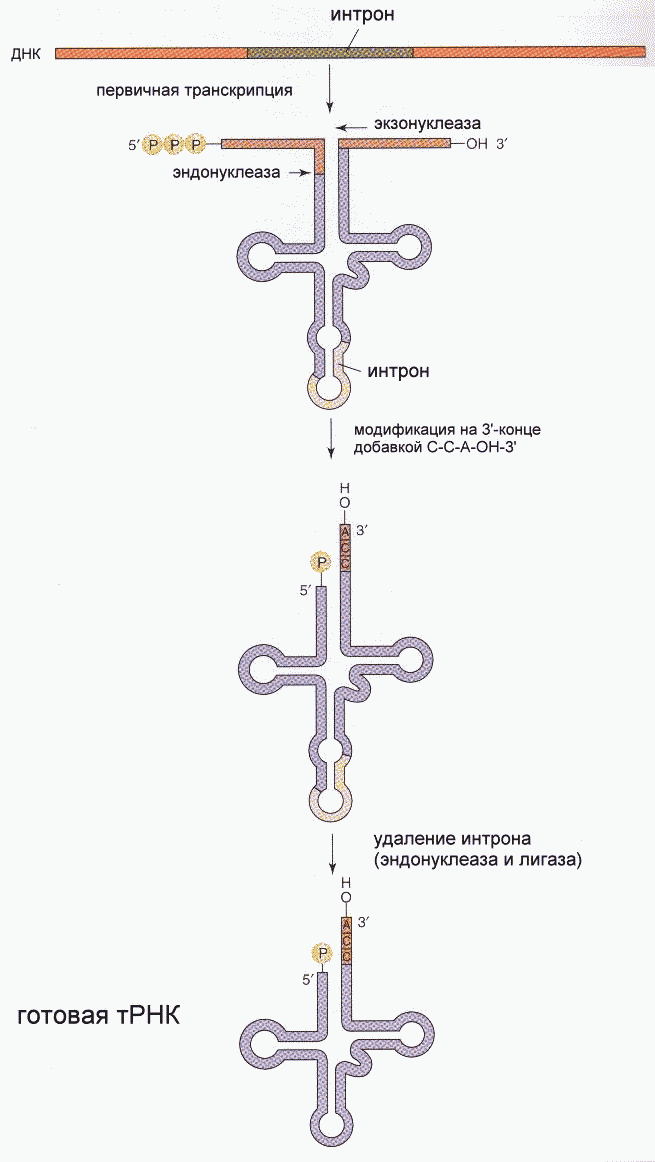
Фрагмент Кленова- результат частичного протеолиза ДНК-полимеразы I E. Coli субтилизином. Основная функция полимеразы III- синтез цепи, полимеразы I- синтез и исправление ошибочно вставленных нуклеотидов. Полимераза II осуществляет особые, спе циализированные функции. Репликация начинается с расплетания цепей ДНК специальными расплетающими белками, которые называют ГЕЛИКАЗАМИ (или Rep-протеином). Геликазы используют энергию АТФ в процессе расплетания цепей. Скорость расплетания составляет около 6000 мин-1. Для того, чтобы расплетенные цепи не могли вновь соединиться, имеются специальные SSB-белки (single-strand binding proteins), которые присоединяются к комплементарным цепям, удерживая их от ассоциации. По мере продвижения репликационной вилки SSB-протеины передвигаются по цепи, диссоциируя с одного места и присоединяясь на другом. Этот процесс не требует затрат энергии АТФ. После освобождения достаточного места начинается синтез праймера- затравки, необходимой для работы ДНК-полимеразы. Наличие затравки является необходимым условием функционирования ДНК-полимераз (как и наличие комплементарной цепи). В качестве затравки на каждой из разделенных цепей синтезируются маленькие отрезки молекул РНК при помощи фермента ПРИМАЗЫ. Синтез новой цепи ДНК осуществляется всегда в направлении 5'-3' , поэтому если по одной матричной цепи возможен непрерывный синтез, то по комплементарной ей цепи синтез осуществляется только участками. Эти участки синтеза называют фрагментами Оказаки. Когда синтез на одном из фрагментов Оказаки достигает праймера другого фрагмента, РНК-овый праймер удаляется имеющейся у полимераз 5'-3' экзонуклеазной активностью и достраивается дезоксирибонуклеотидами. После этого сахарофосфатный остов между фрагментами сшивается ковалентной связью при помощи фермента ДНК-лигазы. Частота возникновения ошибок при репликации и транскрипции НЕ ПРЕВЫШАЕТ 10 -8 -10 -9 , то есть возможна лишь одна ошибка на сотни миллионов нуклеотидов. Такая точность не может быть обеспечена одним только лишь правилом комплементарности нуклеотидов (обеспечивающим точность 1:10000-1:100000). Репликационный аппарат имеет собственные механизмы "поддержания точности" копирования генетической информации. Этими функциями обладают все ДНК-полимеразы. Модель структуры и функциональных участков (на примере ДНК-полимеразы I) показана на рисунке. Она имеет три зоны активности- полимеризующую в направлении 5'-3', и экозонуклеазные в направлениях 5'-3' и 3'-5'. Области активности разделены пространственно. Вперед (по ходу продвижения полимеразы по матричной цепи ДНК) обращена зона 5'-3' экзонуклеазной активности. Она служит для удаления попадающихся на пути РНК-овых праймеров (затравок). Далее идет собственно синтетическая зона и наконец, зона с экзонуклеазной активностью в направлении 3'-5'. С этой зоной связана так называемая PROOF-READING активность (способность узнавать неправильно встроенные нуклеотиды) и исправлять их вырезанием ряда уже встроенных нуклеотидов. Для этого молекула ДНК-полимеразы смещается (не отсоединяясь от ДНК-овой матрицы) к месту синтеза и последовательно вырезает нуклеотиды, после чего возобновляется нормальный синтез. Воздействие на организм неблагоприятных факторов (химические соединения, ультрафиолет и др.) приводит к постоянному накоплению ошибок в геноме, которые, в конечном итоге, вызывают появление патологии, в частности, невыясненный до сих пор механизм раковых заболеваний. Пока лишь существуют только предположения о том, что причиной раковых заболеваний являются дефекты в носителях информации- ДНК. Транскрипция- синтез молекул РНК на основании информации, записанной в ДНК. Осуществляется в ядрах при участии ДНК-зависимых РНК-полимераз, существующих в типах I, II и III (в порядке выхода в гель-хроматографии). В процессе транскрипции копируется не вся информация с ДНК, а только выборочная, часто отрезками. Сигналом для присоединения полимеразы служат так называемые промотеры , в районе которого (35 нуклеотидных пар до и 10 пар после него) и присоединяется РНК-полимераза. Происходит разделение цепей ДНК и начинается синтез молекулы РНК в направлении 5'-3', только на одной из цепей. При этом по месту тиминовых нуклеотидов комплементарной цепи встают уридиловые нуклеотиды. Весь комплекс передвигается по молекуле ДНК, пока не будет закончен синтез требуемого участка РНК. ДНК с "отсканированной" информацией репарирует, ассоциируя в двунитевые молекулы. Возбудитель туберкулеза Micobacterium tuberculosis (точнее, его РНК-полимераза) весьма чувствителен к антибиотику РИФАМПИЦИНУ, в то время как человеческая РНК-полимераза к нему мало чувствительна. На этом свойстве рифампицина основано его использование в терапии туберкулеза. Молекулы РНК очень часто претерпевают посттранскрипционную модификацию, заключающуюся в удалении участков построенной цепи. Наглядно это можно проследить на примере синтеза молекулы транспортной РНК: Обратная транскрипция — создание ДНК-копии по матрице РНК Это правило переноса генетической информации составляет суть молекулярной биологии. Только один раз, в 1970 г., оно было модифицировано, когда признали существование обратной транскрипции. Впервые обратная транскрипция была обнаружена у опухолевых вирусов мышей и кур. Сейчас эти РНК-сод ержашие вирусы называют ретровирусами. Их инфекционный цикл хорошо изучен. Вирус проникает в клетку-мишень, и на основе своей РНК создает копию ДНК, которая встраивается в хромосому хозяина. При делении клетки встроенная копия ДНК вирусного генома удваивается и передается дочерним клеткам. Таким образом, наследственный материал вируса оказывается включенным в геном клетки. Вирусная РНК может образоваться позже путем копирования встроенной ДНК. Это приведет к образованию новых инфекционных вирусов. Именно таким образом вирус иммунодефицита человека (ВИЧ) встраивается в ядерную ДНК и размножается вместе с клеточным геномом. Вот почему от этого вируса трудно избавиться, ведь каждая встроенная (интегрированная) ДНК-копия вируса может использоваться для образования множества копий РНК в результате нормального процесса транскрипции, а это в свою очередь приводит к появлению новых инфекционных ВИЧ-частиц. Еще один пример ретровирусов — это вирусы, вызывающие рак легких и лейкоз у человека и животных. Похожие главы из других книг:Глава 6 ОБРАТНАЯ СВЯЗЬ СОМЫ И ЗАРОДЫШЕВОЙ ЛИНИИ В этой главе мы рассматриваем, как могли изменяться последовательности ДНК вариабельных генов антител в клетках зародышевой линии за эволюционное время. Некоторые данные указывают на существование обратной связи генов, т. Перезапись генетического текста и перевод в белковый текст (транскрипция и трансляция) В клетке ДНК служит в качестве матрицы, на которой первоначально происходит синтез разных РНК. Процесс перезаписи генетической информации из ДНК в РНКовый текст получил название Перезапись генетического текста и перевод в белковый текст (транскрипция и трансляция) В клетке ДНК служит в качестве матрицы, на которой первоначально происходит синтез разных РНК. Процесс перезаписи генетической информации из ДНК в РНКовый текст получил название Целенаправленное создание новых генов В некоторых случаях сомневаться в способности клетки направлять мутационный процесс на нужные гены не приходится. Особенно ярко проявляется это в работе иммунной системы позвоночных. Биологов и медиков давно интересовал вопрос, 7. БИОЛОГИЧЕСКАЯ ОБРАТНАЯ СВЯЗЬ (БОС) Интерес к исследованиям биологической обратной связи (БОС) в целях произвольного управления ФС на основе объективной информации о динамике психофизиологических показателей возник в связи с данными об изменениях в ФС человека, Обратная транскрипция Некоторые РНК-содержащие вирусы (вирус саркомы Рауса, ВИЧ) обладают уникальным ферментом – РНК-зависимой ДНК-полимеразой, часто называемой обратной транскриптазой или ревертазой. Этот фермент обладает время активностями. Первая из них – Транскрипция Транскрипцией называется процесс переноса генетической информации с ДНК на РНК. Матрицей для синтеза РНК служит только одна из двух нитей ДНК (так называемая смысловая цепь). Транскрипция происходит не на всей молекуле ДНК, а на участке одного гена. Ген – Рокфеллер финансирует создание агробизнеса В то время когда братья Рокфеллеры с помощью своего плана Зеленой революции расширяли сферу влияния своего глобального бизнеса в развивающихся странах от нефти до сельского хозяйства, они финансировали малозаметный проект в Создание орудий мышления Как нельзя голыми руками заниматься плотницким делом, так и мышление редко может осуществляться с помощью голого мозга Бо Дальбом, Ларс-Эрик Джанлерт. Компьютерное будущее (в печати). Перед каждым агентом стоит задача наилучшего использования Взаимодействие эндокринных систем: прямая связь, обратная связь, синергизм, пермиссивное действие, антагонизм Как уже было упомянуто, отдельные эндокринные железы и их гормоны взаимодействуют друг с другом, обеспечивая функциональное единство организма. Между Теория главной копии По мнению ведущего молекулярного биолога Харриса Бернштейна (Harris Bernstein) из Аризонского университета, половой процесс возник для починки генов. Первым намеком на это стало исследование особых мутантных дрозофил, не способных репарировать Создание Олиновского нейропсихиатрического исследовательского центра Одним из главных спонсоров Института жизни было семейство Олин, у которого несколько детей лечились в ИЖ. Доктор Хэнк Шварц решил назвать наше новое здание в тюдоровском стиле в их честь. Теперь Создание каталога мира: вербально-концептуальная ассоциативная зона Вербально-концептуальная ассоциативная зона, расположенная в месте пересечения височной, теменной и затылочной долей, в первую очередь отвечает за формирование абстрактных концепций и за их , PhD, Wadsworth Center, NYSDOH Last full review/revision February 2018 by Laura D Kramer, PhD Вирусы – это мельчайшие паразиты, обычно величиной от 0,02 до 0,3 μ м, хотя недавно были обнаружены несколько очень крупных вирусов длиной до 1 μ м (megavirus, pandoravirus). Размножение вирусов находится в полной зависимости от клеток (бактериальных, растительных или животных). Вирусы имеют внешнюю белковую, а иногда и липидную, оболочку, ядро РНК или ДНК и иногда ферменты, необходимые для первых этапов репликации вируса. Вирусы классифицируются преимущественно в соответствии с природой и структурой их генома и способом их репликации, а не в зависимости от заболеваний, которые они вызывают. Таким образом, существуют ДНК-вирусы и РНК-вирусы; каждый тип может иметь одинарные или двойные цепи генетического материала. Одноцепочечные РНК-вирусы в свою очередь подразделяются на РНК с положительной полярностью и РНК с отрицательной полярностью. Как правило, ДНК-вирусы реплицируются в ядре клетки-хозяина, а РНК-вирусы обычно реплицируются в цитоплазме. В то же время, некоторые одноцепочечные РНК-вирусы с положительной полярностью, называемые ретровирусами, используют совершенно другой способ репликации. Ретровирусы используют обратную транскрипцию для создания двухцепочечной ДНК-копии (провируса) их генома РНК, которая встраивается в геном клетки-хозяина. Обратная транскрипция осуществляется с помощью фермента обратной транскриптазы, который вирус несет с собой внутри своей оболочки. Примерами ретровирусов являются вирусы иммунодефицита человека и вирусы Т-клеточного лейкоза человека. После того, как провирус интегрируется в ДНК клетки-хозяина, он транскрибируется с использованием обычных клеточных механизмов для воспроизведения вирусных белков и генетического материала. Если инфицированная клетка относится к зародышевой линии, интегрированный провирус может укорениться как эндогенный ретровирус, который передаётся потомству. Секвенирование генома человека выявило, что как минимум, 1% генома человека состоит из эндогенных ретровирусных последовательностей, представляющие собой предыдущие контакты с ретровирусами в процессе эволюции человека. Немногие эндогенные ретровирусы человека остаются транскрипционно активными и вырабатывают функциональные белки (например, синтицины, которые участвуют в образовании структуры человеческой плаценты). Ряд экспертов полагает, что некоторые заболевания неясной этиологии, такие как рассеянный склероз, определенные аутоиммунные нарушения, а также различные злокачественные опухоли, могут быть вызваны эндогенными ретровирусами. Поскольку транскрипция РНК не использует те же самые механизмы для контроля ошибок, что и транскрипция ДНК, РНК-вирусы – в частности, ретровирусы – особенно склонны к мутации. Для того, чтобы произошло инфицирование, вирус в первую очередь прикрепляется к одиночной или к одной из нескольких рецепторных молекул на поверхности клетки-хозяина. Затем вирусная ДНК или РНК проникает в клетку-хозяин и отделяется от внешней оболочки (декапсуляция вируса) и воспроизводится в клетке-хозяине с участием определенных ферментов. Вновь синтезированные компоненты вируса затем собираются в полноценные вирусные частицы. Клетка–хозяин, как правило, погибает, выделяя новые вирусы, которые заражают другие клетки хозяина. Каждый этап вирусной репликации задействует различные ферменты и субстраты и дает возможность для противодействия процессу инфицирования. Последствия вирусного инфицирования в значительной степени варьируются. Многие инфекции вызывают острое заболевание после непродолжительного инкубационного периода, а некоторые являются бессимптомными или вызывают несущественные симптомы, которые никак нельзя распознать, кроме как ретроспективно. Многие вирусные инфекции устраняются защитными силами организма, но некоторые переходят в латентную форму, а некоторые становятся причиной хронического заболевания. При скрытой инфекции вирусная РНК или ДНК остается в клетках-хозяина, но не реплицируется и не вызывает заболеваний в течение долгого времени, иногда в течение многих лет. Латентные вирусные инфекции могут передаваться в течение бессимптомного периода, что облегчает распространение от человека к человеку. Иногда реактивацию вызывает инициирующий фактор (в частности, иммуносупрессия). К распространённым вирусам, которые переходят в латентную форму, относятся Читайте также:
 Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу. Пожалуйста, не занимайтесь самолечением!При симпотмах заболевания - обратитесь к врачу.
Copyright © Иммунитет и инфекции
|