
Рга для идентификации вирусов

Основными видами диагностики вирусных инфекционных заболеваний, несмотря на разнообразие современных методов лабораторных исследований, остаются серологические тесты и классический метод выделения вирусов на культуре клеток. Успех своевременной этиологической диагностики хантавирусных инфекций в значительной степени зависит от разработки новых и совершенствования рутинных иммунологических методов исследования. Трудности, связанные с выделением на культуре клеток хантавирусов от больных геморрагической лихорадкой с почечным синдромом (ГЛПС) и грызунов-носителей, выдвигают на первое место серологические тесты. Для выявления специфических антител к определенному вирусу чаще всего используют метод непрямой иммунофлюоресценции (НМФА). К вирусам, обладающим гемагглютинирующими свойствами, применяют также и иммунологическую реакцию торможения гемагглютинации (РТГА), которая основана на способности сывороточных антител, вырабатываемых к гемагглютининам (специфическим белкам, содержащимся во внешней оболочке некоторых вирусов), подавлять вызываемую вирусом агглютинацию эритроцитов. После успешной изоляции прототипного штамма 76-118 вируса Hantaan [8] рН-зависимые гемагглютинины были выявлены в антигенах, полученных из мозга новорожденных мышей, клеток VERO E-6, вируссодержащих культуральных жидкостей [6, 10, 11]. Чувствительность и специфичность тестов гемагглютинации в значительной степени зависит от источника и способа получения антигенов, при этом условия репродукции вирусов оказывают заметное влияние на формирование гемагглютининов, а воздействие физических и химических факторов влияет на функциональные свойства антигенов и их стабильность [1, 10]. Было отмечено [9], что тест РТГА по своей специфичности не уступает реакции нейтрализации с использованием тканевых культур, а его применение предпочтительнее в тех регионах, где одновременно циркулируют несколько антигенных вариантов хантавирусов. На территории Приморского края с помощью молекулярно-генетических методов исследования выявлена циркуляция трех патогенных хантавирусов – Hantaan (геновариант FE), Amur и Seoul (геновариант VDV), природными хозяевами для которых установлены восточный подвид полевой мыши (Apodemus agrarius), восточно-азиатская мышь (Apodemus peninsulae) и серая крыса (Rattus norvegicus) соответственно [3–5].
Цель работы – показать эффективность модифицированных тестов торможения гемагглютинации при идентификации штаммов близкородственных вирусов Amur и Hantaan (геновариант FE), изолированных от экологически разных видов мышей рода Apodemus – A. peninsulae и A. agrarius соответственно, а также случаев заболевания ГЛПС, обусловленных этими патогенами, на территории Приморского края.
Материал и методы исследования
Гемагглютинирующие антигены штаммов – прототипных и выделенных нами на клеточной культуре VERO E-6 от грызунов-носителей хантавирусов на территории края – готовили из вируссодержащих культуральных жидкостей по разработанному способу [2]. Исследовали сыворотки крови от инфицированных хантавирусом грызунов (n = 86) и больных ГЛПС (n = 246) из трех очаговых регионов края (I – Восточно-Маньчжурский холмисто-равнинный, II – Амуро-Уссурийский предгорно-лесной, III – Сихотэ-Алиньский горно-таежный [7]) и г. Владивостока. Все больные были с серологически подтвержденным в НМФА диагнозом ГЛПС без четкого различия в титрах антител к вирусам Hantaan, Amur и Seoul. Гемагглютинирующую активность антигенов определяли в реакции гемагглютинации (РГА). Этиологическую диагностику ГЛПС у больных из разных регионов края и в разные сроки заболевания проводили в условиях модифицированной постановки РТГА – 2 активные единицы антигена (АЕ), время контакта 15 минут и 2 часа при +4 °С. Антигенные связи штаммов хантавирусов изучали в предложенной кинетической постановке РТГА (КРТГА) – 4 АЕ антигена, время контакта 15 и 30 минут, 1, 2, 4 и 18 часов при + 4 °С (табл. 1).
Характеристика тестов гемагглютинации, используемых в работе
Реакция гемагглютинации (РГА)
Определение гемагглютинирующей активности антигенов и их рабочей дозы – 4-8 АЕ
Реакция торможения гемагглютинации (РТГА)
Оценка специфической гемагглютининингибирующей активности антител (антигемагглютининов) в сыворотках крови больных ГЛПС по ? 4-кратной разнице в титре антител к антигенам гомо- и гетерологичных хантавирусов
Кинетическая реакция торможения гемагглютинации (КРТГА)
Проведение серологической идентификации штаммов хантавирусов с учетом кинетики взаимодействия антигемагглютининов иммунных сывороток крови, полученных к серотипам Hantaan (HTNV), Amur (AMRV), Seoul (SEOV) и Puumala (PUUV), с гомо- и гетерологичными гемагглютинирующими антигенами хантавирусов
Внутри- и межтиповые антигенные отношения исследуемых штаммов хантавирусов оценивали в перекрестных КРТГА, при этом степень антигенного сходства или различия штаммов количественно определяли по рассчитанному в каждой реакции значению предложенного ранее показателя А [1].
Результаты исследования и их обсуждение
На первом этапе изучали эффективность теста РТГА для дифференциальной диагностики хантавирусных инфекций среди разных видов – носителей хантавирусов. Часть результатов параллельного титрования сывороток крови от инфицированных животных с гемагглютинирующими антигенами разных хантавирусов представлена в табл. 2. Специфические антитела чаще выявляли к тому хантавирусу, для которого данный вид животного является основным хозяином. В ряде случаев отмечали перекрестные реакции с гетерологичными вирусами, но титр антител к гомологичному вирусу был выше.
Идентификация хантавирусной инфекции у грызунов-носителей в РТГА
Номер исследуемой сыворотки
Гемагглютинирующий антиген хантавируса / грызун-носитель
1. Выделение и идентификация респираторных вирусов
Частота выделения респираторных вирусов зависит от сроков взятия материалов от больных, условий их хранения, доставки и времени первичной обработки. Решающее значение имеет ассортимент и качество клеточных культур и куриных эмбрионов, на которых проводят изоляцию вирусов.
Рекомендуемый состав культур для изоляции вирусов гриппа и ОРЗ:
1) первичная культура клеток почек эмбриона человека (ПЭЧ);
2) перевиваемые культуры клеток МДСК, Неla (или HEp2);
3) фибробласты легкого эмбриона человека (ЛЭЧ).
Материалом для выделения вирусов служат отделяемое носа, зева, конъюнктивы, а также секционный материал (ткани легкого, кусочки бронхов и др.).
Материал отбирают с помощью стерильного тампона, который погружают затем в пробирки с 5,0 мл среды Игла или среды 199 с антибиотиками и немедленно направляют в лабораторию, где тампоны с вирусным материалом отжимают в среду, материал центрифугируют в течение 20 мин при 2000-3000 об/мин и температуре 4°С или используют в неосветленном виде.
Кусочки секционного материала растирают в фарфоровой ступке со стеклянным песком, добавляют 10-кратное количество раствора Хенкса, взвесь ресуспендируют, после чего осветляют центрифугированием. Надосадочную жидкость используют для заражения куриных эмбрионов или тканевых культур.
1.1. Изоляция и идентификация вирусов
Для выделения вирусов гриппа исследуемые материалы после их первичной обработки вводят по 0,1 мл в амниотическую полость и по 0,2 мл в аллантоисную полость 10-дневных куриных эмбрионов.
Заражение проводят в затемненном боксе через боковую поверхность скорлупы, направляя иглу сначала в амниотическую, а затем в аллантоисную полости, предварительно сделав дополнительный прокол в скорлупе над воздушной камерой. Материалом от одного больного заражают 3 эмбриона.
Зараженные эмбрионы инкубируют в термостате 72 часа при 33-34°С. После инкубации эмбрионы помещают на ночь в рефрижератор при 4°С.
Индикацию вируса гриппа осуществляют в реакции гемагглютинации (РГА) путем добавления 0,5 мл 1% суспензии отмытых эритроцитов кур к 0,5 мл вируссодержащего материала. Результаты РГА учитывают после 30-минутного оседания эритроцитов в лунках панелей при комнатной температуре.
При отсутствии агглютинации эритроцитов необходимо провести 3 дополнительных пассажа путем заражения эмбрионов смесью амниотической и аллантоисной жидкостей от предыдущего пассажа. В случае отрицательных результатов РГА после 3-х пассажей исследование материалов прекращают.
Идентификацию вирусов гриппа проводят в РТГА.
Для этого к 0,2 мл последовательных разведений типоспецифических иммунных сывороток добавляют по 0,2 мл рабочей дозы вируса в количестве 4 гемагглютинирующих единиц (ГЕ). Одной ГЕ считают последнее разведение вируса, дающее отчетливую гемагглютинацию. После 1-часового контакта сыворотки с вирусом при комнатной температуре в каждую лунку добавляют по 0,4 мл 1% взвести куриных эритроцитов. Титром сывороток считают ее последнее разведение, блокирующее 4 ГЕ вируса.
Типовую принадлежность вируса определяют по ингибиции гемагглютинации, которая должна быть зарегистрирована в разведении не менее, чем 1:20.
Выделение вирусов гриппа может быть осуществлено на чувствительных культурах клеток, чаще всего МДСК. Для этого в пробирки монослоем, отмытым физиологическим фосфатным буфером или раствором Хэнкса, вносят по 0,2 мл материала от больного. Через 1 ч адсорбции при 38 °С в культуры добавляют по 1,5 мл поддерживающей среды (199, Игла с антибиотиками) и помещают в термостат при 33°С.
Индикацию, типирование и дополнительные пассажи вирусов гриппа проводят как указано выше. Кроме того, инфицированные культуры 1 раз в день микроскопируют для оценки цитопатогенного действия вирусов.
Изоляцию аденовирусов, парагриппозных вирусов, РС-вируса, коронавирусов, вирусов герпеса и др. проводят на клеточных культурах (ПЭЧ, ЛЭЧ и Нер-2). Наиболее чувствительные культуры вносят по 0,2 мл материала от больного. Через 30-60 мин адсорбции вируса на клетках в культуры вносят по 0,8 мл поддерживающей среды Игла с антибиотиками (100 ед/мл пенициллина, 100 мкг/мл стрептомицина) и с 2% инактивированной фетальной сыворотки. Зараженные культуры инкубируют при температуре 36 °С до появления в них проявлений цитопатогенного действия (ЦПД).
Индикацию парагриппозных вирусов проводят по ЦПД и гемадсорбции с 0,4% взвесью эритроцитов морской свинки (20 мин при комнатной температуре), а остальных вирусов
- по характерному цитопатогенному действию.
Если при первичном заражении вирус не был выявлен ни в одном из тестов, материалы дополнительно пассируют еще 2 раза в тех же условиях.
Идентификацию вирусов ОРЗ проводят в чувствительных тканевых культурах с помощью реакции нейтрализации (РН) со специфическими иммунными сыворотками. Вируссодержащую культурную жидкость в дозе 100 ТИД50/0,2 мл в объеме 0,2 мл смешивают с 0,2 мл каждой из типоспецифических иммунных сывороток, взятых в разведении 1:20. Смесь вируса и сыворотки выдерживают 2 часа при комнатной температуре, после чего по 0,2 мл каждой смеси вносят в 2 пробирки с монослоем чувствительных клеток, а затем в пробирки добавляют по 0,8 мл поддерживающей среды. Инфицированные и контрольные культуры клеток инкубируют при температуре 36 °С. Параллельно типоспецифические сыворотки контролируют в отношении их активности к типовым эталонным штаммам.
Учет реакции проводят по ингибиции цитопатогенного действия (для аденовирусов, РС, простого герпеса), гемадсорбции (для парагриппозных вирусов) в день, когда типируемый вирус вызвал выраженное ЦПД клеток в контрольных культурах без сыворотки (при отсутствии дегенерации в контрольных посевах неинфицированных клеток и в контролях диагностических сывороток), в то время как эталонные штаммы полностью нейтрализованы типоспецифическими сыворотками. Тип выделенного штамма определяют по сыворотке, которая полностью предохранила клетки от дегенерации.
метод обнаружения и идентификации вирусов, основанный на наличии у некоторых вирусов способности избирательно агглютинировать эритроциты определенных видов животных.
В основе РГА лежит феномен склеивания эритроцитов, происходящий под влиянием различных факторов. Различают прямую и непрямую гемагглютинацию. При реакции прямой гемагглютинации происходит склеивание эритроцитов при адсорбции на них определенных антигенов, например вирусов.
В серологических исследованиях применяют реакцию торможения прямой гемагглютинации, когда выделенный у больного вирус нейтрализуют специфической иммунной сывороткой, а затем соединяют с эритроцитами. Отсутствие гемагглютинации говорит о соответствии вируса и используемой иммунной сыворотки.
Реакция непрямой гемагглютинации (пассивная гемагглютинация) наблюдается в тех случаях, когда к эритроцитам, заранее обработанным (сенсибилизированным) различными антигенами, прибавляют иммунную сыворотку или сыворотку больного, имеющую соответствующие антитела. Происходит специфическое Склеивание эритроцитов, их пассивная гемагглютинация.
Реакция непрямой, или пассивной, гемагглютинации по чувствительности и специфичности превосходит другие серологические методы, и ее используют при диагностике инфекций, вызванных бактериями, риккетсиями, простейшими.
РГА используется для индикации (обнаружения) вирусов при проведении ориентировочной диагностики, для титрования вирусов по гемагглютинирующим свойствам (установление гемагглютинирующих единиц - АЕ).
Основу феномена агглютинации, вызываемого вирусами, составляет адсорбция вирусов на поверхности эритроцитов, сопровождающаяся склеиванием (агглютинацией) последних и выпадением в осадок.
Антиген для РГА
В качестве антигена для РГА берут любой материал (патматериал в виде суспензии из органов, материал из зараженных КЭ, культур ткани и др.), в котором предполагается наличие вируса. Материал должен быть жидким, без крупных частичек.
Постановка ориентировочной РГА
Для постановки ориентировочной РГА на чистое и хорошо обезжиренное предметное стекло наносят одну каплю вирусосодержащего материала, к ней добавляют одну каплю 5% взвеси эритроцитов и перемешивают стеклянной палочкой.
Оценка реакции РГА
Оценивают реакцию в крестах (плюсах). При оценке реакции в крестах обращают внимание на характер осадка. Если эритроциты осели тонким слоем равномерно по дну пробирки (в виде зонтика), реакцию оценивают в четыре креста
2. Реакция торможения гемагглютинации — серологическая реакция, основанная на способности антител предотвращать агглютинацию эритроцитов гемагглютинирующими видами вирусов (аденовирусами, арбовирусами, некоторыми энтеровирусами, вирусами гриппа и парагриппа, кори, реовирусами). Специфические антивирусные антитела взаимодействуют с поверхностными молекулами гемагглютининов вирионов этих вирусов и блокируют их связывание с комплементарными им молекулами мембраны эритроцитов.
В последнее время реакция широко используется в лабораториях клинической вирусологии для определения титров специфических антител к тем или иным вирусам, а также для серологической идентификации и типирования изолятов вирусов из клинического материала от больных. Используют несколько ограничено в силу наличия в сыворотке крови людей неспецифических ингибиторов вирусов, а также естественных антител — агглютининов.
Реакция торможения гемагглютинации
метод идентификации вируса или выявления противовирусных антител в сыворотке крови больного, основанный на феномене отсутствия агглютинации эритроцитов препаратом, содержащим вирус, в присутствии иммунной к нему сыворотки крови.
Реакция торможения гемагглютинации. Механизм и практическое использование.
Многие вирусы обладают способностью агглютинировать эритроциты строго определенных видов млекопитающих и птиц. Так, вирусы гриппа и эпидемического паротита агглютинируют эритроциты кур, морских свинок, человека, а аденовирусы – эритроциты крыс, мышей. В связи с этим для их обнаружения в материале больных или культурах клеток, эмбрионов и животных ставят реакцию гемагглютинации (РГА). Для этого в лунках планшетов готовят двукратно возрастающие разведения вируссодержащих материалов и жидкостей, добавляя к ним отмытые изотоническим раствором взвеси NaCl эритроцитов. Для контроля спонтанной агглютинации эритроциты смешивают ещё с равным объемом изотонического раствора NaCl. Смеси инкубируют в термостате при температуре 37°С или при комнатной температуре.
Являясь группоспецифической, РГА не дает возможности определить видовую принадлежность вирусов. Их идентифицируют с помощью реакции торможения гемагглютинации (РТГА). Для ее постановки используют заведомо известные иммунные противовирусные сыворотки, которые в двукратно снижающихся концентрациях разводят в изотоническом растворе натрия хлорида и разливают по лункам. К каждому их разведению добавляют равное количество вируссодержащей жидкости. Контролем является взвесь вируса в изотоническом растворе натрия хлорида. Планшеты со смесью сывороток и вируса выдерживают в термостате 30 мин или при комнатной температуре 2 ч, затем в каждую из них добавляют взвесь эритроцитов. Спустя 30 мин определяют титр вируснейтрализующей сыворотки (т.е. максимальное ее разведение), вызвавшей задержку агглютинации эритроцитов.
Используют РТГА в серологической диагностике вирусных болезней, в частности гриппа и аденовирусных инфекций. Ставить ее лучше так же, как и РН, с парными сыворотками. Четырехкратное нарастание титра антител во второй сыворотке подтверждает предполагаемый диагноз
3. Серологическое исследование позволяет поставить диагноз в случае обнаружения специфических антител в сыворотке больных. Для серологических реакций используют парные сыворотки, которые берут в первые дни от начала заболевания и спустя 1 - 3 недели, но изучают одновременно. Диагностическое значение имеет нарастание титра антител в 4 раза и более. РТГА, РН, РСК, РТГадс, РИФ, РИА, ИФА ставят с антигенами ( диагностикума-ми), приготовленными из эталонных штаммов соответствующих вирусов.
4. Парные - это значит, что кровь дважды берут с каким-то интервалом времени. По нарастанию титра антител определяют, что заражение произошло. Если титр антител не меняется, значит, эти антитела в организме уже давно, свежего заражения нет.
Наибольшую диагностическую ценность представляет исследование парных сывороток, взятых от животных в начале и в конце заболевания (с промежутком в 14-21 день). Сыворотку отбирают в стерильные пробирки и хранят для исследования.
Набор для диагностики парвовирусной болезни свиней предназначен для обнаружения парвовирусного антигена в суспензии внутренних органов абортированных плодов в реакции гемагглютинации (РГА) и специфических антител в сыворотке крови свиней, а также новорожденных поросят (до приема молозива) в реакции торможения гемагглютинации (РТГА).
Реакцию ставят микрометодом в лунках полистироловых пластин.
полистироловые 96-луночные планшеты для иммунологических реакций;
антиген специфический инактивированный, обладающий гемагглютинирующей активностью не ниже 1:128;
сыворотка специфическая, обладающая активностью в РТГА не ниже 1:256;
сыворотка нормальная (отрицательный контроль).
Время проведения анализа:
РГА – 2,5-3,5 часа, РТГА – 3,5-4,5 часа (не учитывая время подготовки материалов для исследования).
Реакция гемагглютинации (РГА) основана на склеивании взвешенных в жидкости эритроцитов под воздействием гемагглютинирующих свойств вируса и образования рыхлого осадка в виде зонтика. Сущность РТГА заключается в нейтрализации гемагглютинирующих свойств вируса специфическими антителами, содержащимися в сыворотке крови, в результате чего не происходит склеивания эритроцитов и они оседают на дно, образуя плотный осадок в виде пуговки.
Набор хранят в сухом, защищенном от света месте при температуре от 2 до 8°С.
После растворения компоненты набора можно хранить в замороженном состоянии в течение 1 месяца, повторное замораживание не допускается.
Эритроциты (или частицы латекса) с адсорбированными на них антигенами взаимодействуют с соответствующими антителами сыворотки крови, что вызывает склеивание и выпадение эритроцитов на дно пробирки или ячейки в виде фестончатого осадка. При отрицательной реакции эритроциты оседают в виде пуговки.
Реакцию агглютинации можно проводить в микроварианте в ячейках 96-луночного планшета для иммунологических исследований с коническим дном. В лунки планшета вносят по 0,05 мл ФСБ (рН 7,2-7,4) и готовят двукратные разведения испытуемых сывороток крови от 1:2 и выше. Затем в каждую ячейку вносят по 0,005 мл суспензии грибных клеток в концентрации 100 тыс. грибных клеток в 1 мл. Планшет осторожно встряхивают и выдерживают 2 часа в термостате при 37°С, а затем 16-18 часов - при 4°С. В качестве отрицательного контроля используют нормальную (негативную) сыворотку крови и ФСБ.
Результаты реакции учитывают с помощью микроскопа и визуально и определяют в крестах по следующей схеме:
(++++) - полное просветление жидкости и образование агглютината на дне лунки в виде перевернутого "зонтика", при встряхивании "зонтик" разбивается на хлопья;
(+++) - неполное просветление жидкости и хорошо выраженный "зонтик";
(++) - заметное просветление жидкости, "зонтик" выражен умеренно;
(+) - едва заметное просветление жидкости, "зонтик" выражен слабо;
(-) - отрицательный результат, просветление жидкости не наступило, на дне лунки осадок в виде пуговки, при легком встряхивании образуется равномерная взвесь.
За титр антител принимали последнее разведение испытуемой сыворотки, в котором произошла агглютинация не менее чем в два креста (++).
 | Из за большого объема этот материал размещен на нескольких страницах: 1 2 3 |

Гемагглютинация, вызываемая вирусами, не является иммунологической реакцией, так как здесь нет системы антиген – антитело. С помощью этой реакции легко выявить присутствие гемагглютинирующего вируса, но невозможно его идентифицировать. Вирусы, обладающие гемагглютинирующими свойствами, имеют на поверхности гемагглютинины, с помощью которых происходит склеивание эритроцитов. По своей химической природе гемагглютинины являются глико - или липопротеидами.
В процессе взаимодействия вирусов с эритроцитами различают стадии адсорбции, агглютинации и элюции. Адсорбция вирусов на эритроцитах начинается как неспецифическая, а затем переходит в специфическую вследствие сродства гемагглютининов к рецепторам эритроцитов. На одном эритроците адсорбируется множество вирусов, они образуют мостики между эритроцитами, изменяется электростатический заряд эритроцитов и в результате наступает следующая стадия взаимодействия – гемагглютинация, на которой процесс может остановиться. Некоторые вирусы способны к элюции – освобождению с поверхности эритроцитов. Элюция происходит под действием вирусных ферментов (например, нейраминидазы) на рецепторы эритроцитов. Элюированные вирусы при этом не повреждаются, эритроциты же не способны повторно адсорбировать вирус вследствие разрушения рецепторов.
При постановке РГА используют те эритроциты (птиц, животных, человека), с которыми гемагглютинирующие свойства данного вируса проявляются лучше (например, эритроциты кур). Задерживающее действие на РГА могут оказывать ингибиторы, содержащиеся в тканях, где размножался вирус, и других биологических субстратах. Для снятия ингибиторного действия вируссодержащие материалы прогревают, обрабатывают трипсином, ацетоном и др.
2.2.1. Техника постановки РГА
Схема постановки реакции гемагглютинации при диагностике гриппа
Номера лунок разведения вируссодержащего материала
1% взвесь эритроцитов
Если предварительно соединить вирус со специфической иммунной вируснейтрализующей сывороткой и потом добавить эритроциты, то такой инактивированный вирус теряет способность вызывать гемагглютинацию. Эта реакция получила название реакции торможения гемагглютинации (РТГА) и может быть использована для окончательной идентификации неизвестного гемагглютинирующего вируса – по специфической диагностической сыворотке, а также для выявления титра неизвестных антител в сыворотке больного – по известным вирусным антигенам-диагностикумам.
Затрудняет применение РТГА наличие в сыворотках крови человека и животных неспецифических вирусных ингибиторов, которые могут вызвать неспецифическое торможение гемагглютинации. Выпускают противовирусные диагностические сыворотки, освобожденные от ингибиторов. РТГА ставят обычно на досках с лунками по методике, сходной с РГА.
2.2.2. Техника постановки РТГА
РТГА с целью идентификации вируса гриппа рода А. На плексигласовой доске готовят три ряда двукратных возрастающих разведений (в объёме 0,25 мл) диагностических противогриппозных сывороток А(Н1N1), А(Н2N2) и А(Н3N2) от 1:10 до 1:320 (до титра сывороток). Во все лунки, кроме контрольной, добавляют по 0,25 мл рабочей дозы вируса (разведение 1:40), в котором содержится 4 гемагглютинирующие единицы (табл. 5). После выдерживания реакции в течение 1 часа при комнатной температуре во все лунки добавляют по 0,5 мл 1% взвеси эритроцитов кур. Учёт результатов проводят через 30-40 минут. В примере торможение произошло сывороткой А(Н3N2), следовательно выделен вирус гриппа, принадлежащий к подтипу А(Н3N2).
Схема постановки РТГА для определения типовой принадлежности
вируса гриппа А
Номера лунок, разведения иммунной сыворотки
Диагностические противогриппозные сыворотки в
Вируссодержащая аллантоисная жидкость в разведении 1:40
Взвесь эритроцитов кур 1%
Выдерживание 1 час при комнатной температуре
РТГА ставят и для определения наличия и титра антител в сыворотке больного. Для этого готовят последовательные двукратные разведения сыворотки в объёме 0,25 мл и соединяют их с 0,25 мл известного антигена (содержащего 4 гемагглютинирующие единицы), чтобы произошла нейтрализация вируса. Затем в пробирки вносят по 0,5 мл взвеси эритроцитов. За титр сыворотки принимают то наибольшее её разведение, в котором ещё наблюдается торможение гемагглютинации (1:160).
Основными составляющими компонентами этой реакции являются: иммунная вируснейтрализующая (опытная) и нормальная (контрольная) сыворотки одного и того же вида животного или человека; содержащий живые вирусы материал, являющийся антигеном; биологические объекты (мыши, куриные эмбрионы), на которых обнаруживают вируснейтрализующее действие иммунной сыворотки. Применяют два варианта постановки реакции нейтрализации. При одном из них берут постоянную дозу сыворотки и различные разведения вируса, при втором – постоянное разведение вируса и различные разведения сыворотки. Во всех случаях последовательность проведения реакции следующая:
1) предварительный контакт вируссодержащего материала с соответствующей иммунной сывороткой (один из этих компонентов заведомо известен, наличие другого определяется);
2) введение этой смеси в чувствительную биологическую систему (белым мышам или куриным эмбрионам);
3) учёт наличия или отсутствия нейтрализации.
Сыворотки получают либо от больных людей, либо применяют диагностические сыворотки от иммунизированных животных. Перед постановкой реакции сыворотки инактивируют, прогревая при температуре 56-600С.
Антигены (вируссодержащие взвеси) получают из материала от больных вирусными инфекциями, готовят из органов и тканей заражённых вирусом животных, куриных эмбрионов, а также из инфицированных клеточных культур и подвергают очистке. Перед употреблением антиген титруют для определения его активности.
Биологические объекты выбирают с учётом свойств вируса, чувствительности самого объекта к действию вируса, а также задач и условий исследования.
Постановка реакции нейтрализации (I вариант)
Ставят для идентификации выделенного вируса. Для постановки реакции берут два ряда пробирок, из которых первый ряд является опытным, а второй – контрольным. В первый ряд пробирок наливают определённое количество неразведённой диагностической иммунной сыворотки, во второй – контрольный – такое же количество неразведённой нормальной сыворотки того же вида животного. Отдельно в пробирках готовят несколько разведений вируса, чтобы при добавлении их в пробирки опытного, контрольного ряда получились последовательные 10-кратные разведения вируса (например, 10-1, 10-2, 10-3 и т. д.). Сыворотки и добавляемые разведения вируса должны быть в одинаковых объёмах, например по 0,2 мл. Пробирки с приготовленной смесью ставят в термостат при температуре 370С обычно на 1-2 часа. После этого опытные и контрольные смеси вводят мышам или в куриные эмбрионы. Каждым разведением для достоверности опыта заражают не менее 4 мышей или куриных эмбрионов.
За заражёнными животными (опытными и контрольными) устанавливают наблюдение в течение 2-х недель, регистрируют заболевших и погибших мышей. Учёт реакции проводят путём сопоставления количества погибших опытных и контрольных мышей от соответствующего разведения вируса. Определяют 50%-ную смертельную дозу – LD50, проводя подсчёт по Риду и Менчу. Высчитывают индеек нейтрализации (ИН) по формуле, определяющей количество доз, нейтрализуемых сывороткой:
Титр вируса в опыте с нормальной сывороткой
Титр вируса в опыте с иммунной сывороткой
ИН менее 10 – отрицательный;
ИН – 11-49 – сомнительный;
ИН равный 50 и выше – положительный.
Заражённые эмбрионы инкубируют при температуре 35-370С от 48 до 72 ч. Затем яйца вскрывают, аллантоисную или амниотическую жидкость отсасывают и в ней определяют наличие вируса при помощи реакции гемагглютинации. Если гемагглютинация наступила в контроле (смесь нормальной сыворотки с разведениями вируса) и отсутствует в опыте (смесь иммунной сыворотки с разведениями вируса), то сыворотка обладает нейтрализующим действием, Конечный титр вируса в контроле и в опыте определяют путём статистической обработки.
Постановка реакции нейтрализации (II вариант)
Реакцию ставят с различными разведениями исследуемой сыворотки и постоянной дозой известного живого вируса. Для этого к двукратным возрастающим разведениям сыворотки добавляют равные объёмы определённого разведения вируса (рекомендуется брать 100 или 1000 LD50 вируса). Далее поступают как и в I варианте, то есть выдерживают опытные и контрольные смеси 1-2 часа в термостате, после чего вводят их мышам или куриным эмбрионам.
При постановке реакции нейтрализации на мышах за титр изучаемой сыворотки принимают разведение, дающее защитный эффект у 50% мышей. В реакции на куриных эмбрионах титром сыворотки считается то наибольшее её разведение, которое ещё способно задержать гемагглютинирующее действие вирусов у 50% заражённых куриных эмбрионов.
2.3. Молекулярно-генетические методы исследования в диагностике вирусных инфекций
К таким методам относятся молекулярная гибридизация и полимеразная цепная реакция, которые позволяют обнаружить присутствие в исследуемом материале даже единичных копий генов определяемых вирусов (ДНК или РНК с определённой нуклеотидной последовательностью), и тем самым доказать наличие соответствующей инфекции.
2.3.1. Молекулярная гибридизация (метод молекулярных зондов)
Метод основан на способности двухспиральной ДНК к денатурации и ренатурации. Денатурация – это расхождение цепей при нагревании ДНК до 80-1000С или обработке её щёлочью. Ренатурация – воссоединение цепей с помощью водородных связей при снижении температуры до 40-600С (отжиг) и приобретение ДНК первоначальной двухспиральной структуры. Разъединённые цепи ДНК способны к гибридизации с фрагментами других односпиральных ДНК, имеющих комплементарные участки расположения нуклеотидов. К гибридизации комплементарных нитей способна также РНК, образуя комплексы ДНК – РНК или РНК – ДНК.
Необходимые для молекулярной гибридизации фрагменты ДНК или РНК, с помощью которых выявляют наличие в исследуемом материале комплементарных нитей нуклеиновой кислоты, называются молекулярными зондами. Молекулярные зонды готовят из нуклеиновых кислот, выделенных из различных вирусов, иногда используют вирусную и-РНК, но чаще зондом служит клонированная рекомбинантная ДНК. Имеются наборы молекулярных зондов для определения многих вирусов.
При постановке реакции молекулярной гибридизации зонды метят радиоактивной (Р32), флюоресцентной или биотиновой меткой, соединяют с исследуемым материалом, содержащим определяемую нуклеиновую кислоту, подвергающуюся денатурации. Если зонд комплементарен цепи определяемой нуклеиновой кислоты, происходит гибридизация в комплементарном участке.
После отжига зонд оказывается включённым в ренатурированную нуклеиновую кислоту и может быть обнаружен по имеющейся метке. Выявление наступившей молекулярной гибридизации позволяет установить природу определяемого вируса. Чувствительность этого метода составляет 10-14 г/мл.
2.3.2. Полимеразная цепная реакция (ПЦР) или локальная
ПЦР, как и молекулярная гибридизация, основана на способности ДНК к денатурации и ренатурации и на комплементарности цепей ДНК. Важным принципом реакции является использование термостабильной ДНК-полимеразы, при участии которой происходит амплификация – умножение определяемых генов или фрагментов с определённой нуклеотидной последовательностью ДНК. В результате реакции исследуемый генетический материал накапливается в значительном количестве и может быть легко выявлен и идентифицирован. Чувствительность этой реакции составляет 10-18 г/мл.
В реакции участвуют следующие ингредиенты:
· определяемая ДНК вирусов в испытуемом биологическом материале, который предварительно концентрируется;
· праймеры 2-х типов (олигонуклеотиды) – короткие цепочки ДНК с нуклеотидной последовательностью комплементарной 3,-концам каждой из двух цепей определяемой ДНК. Праймеры получают из нуклеиновых кислот различных вирусов, их нуклеотидную последовательность определяют методом секвенирования;
· свободные нуклеитиды (дезоксирибонуклеозидтрифосфаты 4-х типов с различными азотистыми основаниями) – необходимый материал для осуществления амплификации;
· фермент термостабильная ДНК-полимераза – производит достройку комплементарных цепей ДНК из пула свободных нуклеотидов. Фермент выделяют из бактерий Thermus aquaticus или получают генно-инженерным методом.
В результате одного цикла ПЦР количество молекул определяемой ДНК удваивается, то есть происходит амплификация ДНК. Обычно проводят 25-40 повторных циклов амплификации и в итоге за 2-3 часа получают миллионы копий специфического фрагмента ДНК вирусов.
ПЦР проводят в 0,5-1,5 мл микроцентрифужных пробирках в амплификаторах, которые автоматически регулируют смену температуры. Каждому из 3-х этапов цикла амплификации – денатурации ДНК, отжига и достройки – необходима инкубация образцов при различном температурном режиме.
1. Денатурация – разъединение определяемой двухспиральной ДНК на две изолированные цепи при нагревании до 90-950С в течение 0,5-1,0 мин.
2. Отжиг – восстановление двухцепочечной структуры определяемой ДНК в области присоединения комплементарного праймера – проводится при температуре 40-600С 0,5 мин.
3. Удлинение (элонгация) – достройка каждой цепи определяемой ДНК доисходного двухцепочечного состояния с помощью термостабильной ДНК-полимеразы – проводится при температуре 70-750С в течение 2-5 мин.
Наличие ДНК после повторных циклов амплификации определяют с помощью электрофореза в полиакриламидном геле, авторадиографией или другими методами.
С помощью ПЦР возможно определение не только нуклеотидной последовательности ДНК, но также РНК и, следовательно, выявление РНК-вирусов (для этого в реакцию вводят обратную транскриптазу). ПЦР особенно ценен для диагностики латентных вирусных инфекций и ВИЧ-инфекции.
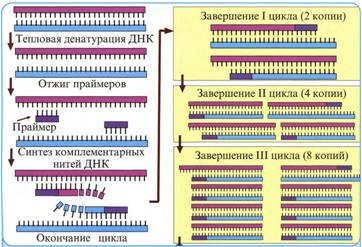
Рис. 8. Схема полимеразной цепной реакции
1. Индикация вируса по его цитопатическим действиям (ЦПД) и определение титра вируса. Идентификация вируса по нейтрализации ЦПД.
2. Метод гемадсорбции, его практическое применение, идентификация вируса в реакции задержки гемадсорбции.
3. Метод бляшек. Идентификация вирусов методом бляшек, титрование антител.
4. Метод цветной пробы, методика постановки, практическое применение.
5. Реакция нейтрализации по цветной пробе, её сущность, применение.
6. Реакция гемагглютинации (РГА), вызываемая вирусами. Практическое применение РГА. Методика постановки.
7. Сущность реакции торможения гемагглютинации (РТГА). Методика постановки. Практическое применение.
8. Реакция нейтрализации вирусов in vivo, способы постановки. Практическое применение.
9. Молекулярно-генетические методы исследования, их сущность. Принцип метода молекулярной гибридизации.
10. Полимеразная цепная реакция (ПЦР). Ингредиенты реакции, их характеристика, значение амплификации. Сущность ПЦР.
11. Техника постановки ПЦР, практическое применение и оценка, как экспресс-метода диагностики вирусных инфекций.
1. Современные методы диагностики распространённых вирусных инфекций.
2. Индикация и идентификация вирусов гриппа.
3. Молекулярно-генетические методы анализа полиморфизма генов.
4. Молекулярно-генетические методы анализа экспрессии генов.
1. Биргер по микробиологическим и вирусологическим методам исследования. 3-е изд. – М.: Медицина, 1982.
2. Большой словарь медицинских терминов / Сост. – М.: , 2007.
3. Вирусология. Методы / Под ред. Б. Мейхи. – М.: Мир, 1988.
4. А., , Широбоков и санитарная микробиология. – М.: АСАDEMA, 2003.
5. Голубев по применению клеточных культур в вирусологии. – Л., 1986. Электронный учебник.
Раздел 1. Культивирование вирусов ……………………………………………4
1.1. Подготовка материала для вирусологических исследований…………….4
1.1.1. Правила работы в вирусологической лаборатории…………………. 4
1.1.2. Материалы, исследуемые при вирусных инфекциях……………. 5
1.1.3. Обработка вируссодержащего материала………………………………5
1.1.4. Микроскопические методы исследования в вирусологии…………….6
1.2. Культуры клеток в вирусологии и методы их получения………………..9
1.2.2. Типы клеточных культур………………………………………………12
1.3. Культивирование вирусов в развивающихся куриных эмбрионах……. 16
1.3.1. Строение куриного эмбриона………………………………………….17
1.3.2. Заражение куриного эмбриона на хорионаллантоисную оболочку. 18
1.3.3. Заражение в аллантоисную полость……………………………. 20
1.3.4. Заражение в полость амниона………………………………………….20
1.4. Культивирование вирусов путём заражения лабораторных животных…22
1.4.1. Интраназальное заражение………………….………………………. 23
1.4.2. Интрацеребральное заражение…………………………………. 24
Раздел 2. Индикация и идентификация вирусов………………………………26
2.1. Методы индикации и идентификации вирусов в клеточных культурах..26
2.1.1. Цитопатическое действие (ЦПД) вируса в культуре клеток……. 27
2.2. Применение реакции гемагглютинации (РГА), реакции
торможения гемагглютинации (РТГА) и биологических
моделей для индикации и идентификации вирусов……………………. 35
2.2.1. Техника постановки РГА………………………………………………36
2.2.2. Техника постановки РТГА……………………………………………..37
2.2.3. Реакция нейтрализации вирусов in vivo………………………………39
2.3. Молекулярно-генетические методы исследования
в диагностике вирусных инфекций……………………………………..41
2.3.1. Молекулярная гибридизация (метод молекулярных зондов)………42
2.3.2. Полимеразная цепная реакция (ПЦР) или локальная
амплификация нуклеиновых кислот (ЛАНК)……………………. 43
Читайте также:
