
Размножение вирусов в микробиологии
Мы уже говорили, что, как только вирусы встречаются с чувствительными клетками, они как бы оживают и начинают вести себя крайне агрессивно. Исследователи, занимавшиеся изучением их размножения, убедились в том, что процесс этот принципиально отличен от известных способов размножения бактерий, простейших и других клеток. Не так давно было установлено, что для размножения вируса необходимо, чтобы вирусная нуклеиновая кислота, которая осуществляет синтез нового вируса, проникла внутрь клеток. Это было обнаружено и экспериментально доказано с помощью остроумных опытов.
В 1956 году удалось разделить вирус табачной мозаики на составляющие его белок и нуклеиновую кислоту. Оказалось, что вирусный белок никак не действует на растения, но после заражения табака изолированной нуклеиновой кислотой отмечалось образование вируса, обладающего всеми свойствами исходного типа. Способность изолированной вирусной нуклеиновой кислоты вызывать заболевание клеток была обнаружена и у возбудителей других вирусных инфекций — полиомиелита, энцефалита, энцефаломиелита, ящура.
Роль РНК и ДНК в передаче наследственности подтвердили и другие факты. Так, после обработки специальными ферментами, разрушающими нуклеиновую кислоту, вирус переставал размножаться. А если на него или на его изолированную нуклеиновую кислоту действовали различными физическими или химическими факторами, изменявшими ее структуру, менялись наследственные свойства и потомства вируса. Итак, размножением вирусов управляют нуклеиновые кислоты.
Не успели ученые ответить на один вопрос, как возникли десятки новых. Как вирусы проникают в клетки? Как из одной вирусной частицы образуются сотни новых? Что происходит внутри зараженных клеток?
Все эти вопросы касаются процесса размножения вирусов. Исчерпывающего ответа на них еще нет, но многое уже ясно.
Атака на клетку начинается с прикрепления вирусных частиц к клеточной стенке, или с так называемой адсорбции вируса. Адсорбция наблюдается сразу же после внесения возбудителей в среду, где есть клетки. Нужно сказать, что каждый вирус способен адсорбироваться лишь на определенных клетках. Считается, что в клеточной стенке есть такие участки — рецепторные поля, которые и соединяются с вирусами.
После прикрепления к клеточной стенке начинается внедрение вирусов в клетку.
Другие вирусы не столь агрессивны. Как это ни странно, им помогает сама клетка. Она как бы заглатывает прикрепленные к ней вирусные частицы. Здесь мы встречаемся с тем случаем, когда созданное многовековой эволюцией целесообразное приспособление приводит к нежелательным результатам.
Посудите сами, активное поглощение клетками из окружающей среды различных частиц (фагоцитоз) или капелек воды необходимо для нормальной жизнедеятельности клетки, а использование этого же механизма для захвата вирусов скорее напоминает самоубийство.
Некоторые наиболее сложно устроенные вирусы проникают в клетку с помощью ферментов. Как же это происходит? Клетка живого организма покрыта оболочкой, которая состоит из двух слоев, имеющих различный химический состав. Чтобы попасть внутрь клетки, вирус должен преодолеть оба этих барьера, Для этого у некоторых из них есть специальные ферменты, которые называют входными.
Вирусная нуклеиновая кислота остается в цитоплазме или по клеточным каналам очень быстро проникает в ядро и ядрышки клетки. С этого момента и начинается размножение вируса.
В настоящее время никто не сомневается, что в размножении вирусов ведущая роль принадлежит нуклеиновой кислоте. А играет ли какую-либо роль в этом процессе белок? Для выяснения этого вопроса в качестве модели были использованы бактериофаги. Тщательные исследования позволили обнаружить, что незначительное количество (около 2%) белка локализовано внутри вирусной частицы и, по-видимому, связано с ДНК. Этот белок проникает вместе с ДНК в клетку. Однако его роль в размножении до сих пор непонятна. Предполагают, что этот белок является как бы посредником между ДНК родительского фага и его потомками.
Теперь мы переходим к самому сложному этапу, который нельзя увидеть под микроскопом,— собственно размножению вирусов. Здесь пока много предположений и гипотез, но нет еще цельной, стройной теории. Для этого есть много причин: трудность работы с таким неуловимым объектом, как вирусы, различный характер их размножения, а также несовершенство существующих методов.
В течение этого периода вирус нельзя обнаружить никакими методами, и о его размножении судят лишь по косвенным признакам. Помогли в этом деле меченые атомы. Ввести радиоактивный изотоп в состав вирусной частицы сравнительно несложно. Для этого культуру ткани предварительно выращивают на среде, содержащей изотопы, а затем заражают вирусом. В процессе своего размножения он поглощает часть изотопов и становится меченым. К вирусу, таким образом, как бы подвешивается крохотный передатчик, который все время сигнализирует о его местонахождении.
Исследователи научились вводить изотопы в различные части вируса. Используя радиоактивный фосфор, можно получить вирус с меченой нуклеиновой кислотой; с помощью радиоактивной серы получают вирус, содержащий изотоп в белковой оболочке. Таким образом, с помощью радиоактивных изотопов можно изучить роль отдельных компонентов вируса при его взаимодействии с клетками.
Благодаря меченым атомам удалось установить, что для синтеза различных компонентов вирусной частицы используются соединения, имеющиеся в клетке, и вещества окружающей питательной среды. Предполагается, что вирусы приостанавливают нормальный клеточный обмен веществ и перестраивают уже существующие в клетке механизмы синтеза белка и нуклеиновых кислот, направляя их на производство вирусного белка и вирусной нуклеиновой кислоты.
Вирусы могут существовать в двух формах: внеклеточной (покоящейся) и внутриклеточной (размножающейся).
Размножение вирусов.
Вирусы являются внутриклеточными паразитами и размножаются только в живых клетках. Размножение вирусов резко отличается от размножения других микроорганизмов.
Лекция № 5. Общая вирусология. Классификация, структура и особенности биологии вирусов. Бактериофаги.
Открытие вирусов Д.И.Ивановским в 1892г. положило начало развитию науки вирусологии. Более быстрому ее развитию способствовали: изобретение электронного микроскопа, разработка метода культивирования микроорганизмов в культурах клеток.
Слово “вирус” в переводе с латинского- яд (животного происхождения). Этот термин применяют для обозначения уникальных представителей живой природы, не имеющих клеточного (эукариотического или прокариотического) строения и обладающих облигатным внутриклеточным паразитизмом, т.е. которые не могут жить без клетки.
В настоящее время вирусология- бурно развивающаяся наука, что связано с рядом причин:
- ведущей ролью вирусов в инфекционной патологии человека (примеры- вирус гриппа, ВИЧ- вирус иммунодефицита человека, цитомегаловирус и другие герпесвирусы) на фоне практически полного отсутствия средств специфической химиотерапии;
- использованием вирусов для решения многих фундаментальных вопросов биологии и генетики.
Основные свойства вирусов (и плазмид), по которым они отличаются от остального живого мира.
1.Ультрамикроскопические размеры (измеряются в нанометрах). Крупные вирусы (вирус оспы) могут достигать размеров 300 нм, мелкие- от 20 до 40 нм. 1мм=1000мкм, 1мкм=1000нм.
2.Вирусы содержат нуклеиновую кислоту только одного типа- или ДНК (ДНК- вирусы) или РНК (РНК- вирусы). У всех остальных организмов геном представлен ДНК, в них содержится как ДНК, так и РНК.
3.Вирусы не способны к росту и бинарному делению.
4.Вирусы размножаются путем воспроизводства себя в инфицированной клетке хозяина за счет собственной геномной нуклеиновой кислоты.
5.У вирусов нет собственных систем мобилизации энергии и белок- синтензирующих систем, в связи с чем вирусы являются абсолютными внутриклеточными паразитами.
6.Средой обитания вирусов являются живые клетки- бактерии (это вирусы бактерий или бактериофаги), клетки растений, животных и человека.
Все вирусы существуют в двух качественно разных формах: внеклеточной- вирион и внутриклеточной- вирус. Таксономия этих представителей микромира основана на характеристике вирионов- конечной фазы развития вирусов.
Строение (морфология) вирусов.
1.Геном вирусов образуют нуклеиновые кислоты, представленные одноцепочечными молекулами РНК (у большинства РНК- вирусов) или двухцепочечными молекулами ДНК (у большинства ДНК- вирусов).
2.Капсид - белковая оболочка, в которую упакована геномная нуклеиновая кислота. Капсид состоит из идентичных белковых субъединиц- капсомеров. Существуют два способа упаковки капсомеров в капсид- спиральный (спиральные вирусы) и кубический (сферические вирусы).
При спиральной симметрии белковые субъединицы располагаются по спирали, а между ними, также по спирали, уложена геномная нуклеиновая кислота (нитевидные вирусы). При кубическом типе симметрии вирионы могут быть в виде многогранников, чаще всего- двадцатигранники - икосаэдры.
3.Просто устроенные вирусы имеют только нуклеокапсид, т.е. комплекс генома с капсидом и называются “голыми”.
4. У других вирусов поверх капсида есть дополнительная мембраноподобная оболочка, приобретаемая вирусом в момент выхода из клетки хозяина- суперкапсид. Такие вирусы называют “одетыми”.
Кроме вирусов, имеются еще более просто устроенные формы способных передаваться агентов - плазмиды, вироиды и прионы.
Основные этапы взаимодействия вируса с клеткой хозяина.
1.Адсорбция- пусковой механизм, связанный со взаимодействием специфических рецепторов вируса и хозяина (у вируса гриппа- гемагглютинин, у вируса иммунодефицита человека- гликопротеин gp 120).
2.Проникновение- путем слияния суперкапсида с мембраной клетки или путем эндоцитоза (пиноцитоза).
3.Освобождение нуклеиновых кислот- “раздевание” нуклеокапсида и активация нуклеиновой кислоты.
4.Синтез нуклеиновых кислот и вирусных белков, т.е. подчинение систем клетки хозяина и их работа на воспроизводство вируса.
5.Сборка вирионов- ассоциация реплицированных копий вирусной нуклеиновой кислоты с капсидным белком.
6.Выход вирусных частиц из клетки, приобретения суперкапсида оболочечными вирусами.
Исходы взаимодействия вирусов с клеткой хозяина.
1.Абортивный процесс- когда клетки освобождаются от вируса:
- при инфицировании дефектным вирусом, для репликации которого нужен вирус- помощник, самостоятельная репликация этих вирусов невозможна ( так называемые вирусоиды). Например, вирус дельта (D) гепатита может реплицироваться только при наличии вируса гепатита B, его Hbs - антигена, аденоассоциированный вирус- в присутствии аденовируса);
- при инфицировании вирусом генетически нечувствительных к нему клеток;
- при заражении чувствительных клеток вирусом в неразрешающих условиях.
2.Продуктивный процесс- репликация (продукция) вирусов:
- гибель (лизис) клеток (цитопатический эффект)- результат интенсивного размножения и формирования большого количества вирусных частиц - характерный результат продуктивного процесса, вызванного вирусами с высокой цитопатогенностью. Цитопатический эффект действия на клеточные культуры для многих вирусов носит достаточно узнаваемый специфический характер;
- стабильное взаимодействие, не приводящее к гибели клетки (персистирующие и латентные инфекции) - так называемая вирусная трансформация клетки.
3.Интегративный процесс- интеграция вирусного генома с геномом клетки хозяина. Это особый вариант продуктивного процесса по типу стабильного взаимодействия. Вирус реплицируется вместе с геномом клетки хозяина и может длительно находиться в латентном состоянии. Встраиваться в ДНК- геном хозяина могут только ДНК- вирусы (принцип “ДНК- в ДНК”). Единственные РНК- вирусы, способные интегрироваться в геном клетки хозяина- ретровирусы, имеют для этого специальный механизм. Особенность их репродукции- синтез ДНК провируса на основе геномной РНК с помощью фермента обратной транскриптазы с последующим встраиванием ДНК в геном хозяина.
Основные методы культивирования вирусов.
1.В организме лабораторных животных.
2.В куриных эмбрионах.
3.В клеточных культурах - основной метод.
Типы клеточных культур.
1.Первичные (трипсинизированные) культуры- фибробласты эмбриона курицы (ФЭК), человека (ФЭЧ), клетки почки различных животных и т.д. Первичные культуры получают из клеток различных тканей чаще путем их размельчения и трипсинизации, используют однократно, т.е. постоянно необходимо иметь соответствующие органы или ткани.
2.Линии диплоидных клеток пригодны к повторному диспергированию и росту, как правило не более 20 пассажей (теряют исходные свойства).
3.Перевиваемые линии (гетероплоидные культуры), способны к многократному диспергированию и перевиванию, т.е. к многократным пассажам, наиболее удобны в вирусологической работе- например, линии опухолевых клеток Hela, Hep и др.
Специальные питательные среды для культур клеток.
Используются разнообразные синтетические вирусологические питательные среды сложного состава, включающие большой набор различных факторов роста- среда 199, Игла, раствор Хэнкса, гидролизат лактальбумина. В среды добавляют стабилизаторы рН (Hepes), различные в видовом отношении сыворотки крови (наиболее эффективной считают эмбриональную телячью сыворотку), L-цистеин и L-глютамин.
В зависимости от функционального использования среды могут быть ростовые (с большим содержанием сыворотки крови) - их используют для выращивания клеточных культур до внесения вирусных проб, и поддерживающие (с меньшим содержанием сыворотки или ее отсутствием)- для содержания инфицированных вирусом клеточных культур.
Выявляемые проявления вирусной инфекции клеточных культур.
2.Выявление телец включений.
3. Выявление вирусов методом флюоресцирующих антител (МФА), электронной микроскопией, авторадиографией.
4.Цветная проба. Обычный цвет используемых культуральных сред, содержащих в качестве индикатора рН феноловый красный, при оптимальных для клеток условиях культивирования (рН около 7,2)- красный. Размножение клеток меняет рН и соответственно- цвет среды с красного на желтый за счет смещения рН в кислую сторону. При размножении в клеточных культурах вирусов происходит лизис клеток, изменения рН и цвета среды не происходит.
5.Выявление гемагглютинина вирусов- гемадсорбция, гемагглютинация.
6.Метод бляшек (бляшкообразования). В результате цитолитического действия многих вирусов на клеточные культуры образуются зоны массовой гибели клеток. Выявляют бляшки- вирусные “ клеточно- негативные” колонии.
Название семейства вирусов заканчивается на “viridae”, рода- “virus”, для вида обычно используют специальные названия, например - вирус краснухи, вирус иммунодефицита человека- ВИЧ, вирус парагриппа человека типа 1 и т.д.
Вирусы бактерий (бактериофаги).
Естественной средой обитания фагов является бактериальная клетка, поэтому фаги распространены повсеместно (например, в сточных водах). Фагам присущи биологические особенности, свойственные и другим вирусам.
Наиболее морфологически распространенный тип фагов характеризуется наличием головки- икосаэдра, отростка (хвоста) со спиральной симметрией (часто имеет полый стержень и сократительный чехол), шипов и отростков (нитей), т.е. внешне несколько напоминают сперматозоид.
Взаимодействие фагов с клеткой (бактерией) строго специфично, т.е. бактериофаги способны инфицировать только определенные виды и фаготипы бактерий.
Основные этапы взаимодействия фагов и бактерий.
1.Адсорбция (взаимодействие специфических рецепторов).
2.Внедрение вирусной ДНК (инъекция фага) осуществляется за счет лизирования веществами типа лизоцима участка клеточной стенки, сокращения чехла, вталкивания стержня хвоста через цитоплазматическую мембрану в клетку, впрыскивание ДНК в цитоплазму.
4.Выход дочерних популяций.
Основные свойства фагов.
Различают вирулентные фаги, способные вызвать продуктивную форму процесса, и умеренные фаги, вызывающие редуктивную фаговую инфекцию (редукцию фага). В последнем случае геном фага в клетке не не реплицируется, а внедряется (интегрируется) в хромосому клетки хозяина (ДНК в ДНК), фаг превращается в профаг. Этот процесс получил название лизогении. Если в результате внедрения фага в хромосому бактериальной клетки она приобретает новые наследуемые признаки, такую форму изменчивости бактерий называют лизогенной (фаговой) конверсией. Бактериальную клетку, несущую в своем геноме профаг, называют лизогенной, поскольку профаг при нарушении синтеза особого белка- репрессора может перейти в литический цикл развития, вызвать продуктивную инфекцию с лизисом бактерии.
Умеренные фаги имеют важное значение в обмене генетическим материалом между бактериями- в трансдукции (одна из форм генетического обмена). Например, способностью вырабатывать экзотоксин обладают только возбудитель дифтерии, в хромосому которого интегрирован умеренный профаг, несущий оперон tox, отвечающий за синтез дифтерийного экзотоксина. Умеренный фаг tox вызывает лизогенную конверсию нетоксигенной дифтерийной палочки в токсигенную.
По спектру действия на бактерии фаги разделяют на :
- поливалентные (лизируют близкородственные бактерии, например сальмонеллы);
- моновалентные (лизируют бактерии одного вида);
- типоспецифические (лизируют только определенные фаговары возбудителя).
На плотных средах фаги обнаруживают чаще с помощью спот (spot) - теста (образование негативного пятна при росте колоний) или методом агаровых слоев (титрования по Грациа).
Практическое использование бактериофагов.
1.Для идентификации (определение фаготипа).
2.Для фагопрофилактики (купирование вспышек).
3.Для фаготерапии (лечение дисбактериозов).
4.Для оценки санитарного состояния окружающей среды и эпидемиологического анализа.
Вирусы — группа ультрамикроскопических облигатных внутриклеточных паразитов, способных размножаться только в клетках живых организмов (многоклеточных и одноклеточных). Среди них имеются возбудители заболеваний человека, животных, растений, насекомых, простейших и микроорганизмов.
Вирусы были открыты в 1892 г. Д. И. Ивановским при изучении причин гибели табака от мозаичной болезни, выражающейся в появлении пятен на листьях растений. Ученый обнаружил, что здоровое растение получает возбудителя с соком больного растения даже после пропускания этого сока через бактериологические фильтры. Следовательно, болезнь вызывает организм, который способен проходить через бактериологические фильтры. Эти микроорганизмы назвали фильтрующимися вирусами, а затем просто вирусами.
Вирусы обладают следующими характерными особенностями, отличающими их от других микроорганизмов:
- • не имеют клеточного строения;
- • не способны к росту и бинарному делению;
- • не имеют собственных систем метаболизма;
- • содержат нуклеиновые кислоты только одного типа — ДНК или РНК;
- • используют рибосомы клетки-хозяина для образования собственных белков;
- • не размножаются на искусственных питательных средах и могут существовать только в организме восприимчивого к ним хозяина.
Обычно вирусы существуют в двух формах — внеклеточной в виде так называемого вириона и внутриклеточной, называемой репродуцирующимся, или вегетативным, вирусом. У вириона отсутствует обмен веществ, он не растет и не размножается. Внутриклеточная форма представляет собой активный агент, который, попав в клетку хозяина (растения, животного, микроорганизма), использует ее биосинтетический и энергетический аппарат для репродукции новых вирусов, а впоследствии может вызвать и гибель самой клетки. Следовательно, только в клетке хозяина вирус способен функционировать и репродуцироваться, приобретая свойства живого организма.
Химический состав вирусов довольно прост. Число химических соединений, из которых они состоят, невелико. Вирусы представляют собой нуклеоп роте иды и состоят из нуклеиновой кислоты и нескольких кодируемых ею белков. Нуклеиновые кислоты вирусов отличаются значительным разнообразием, превосходя в этом отношении даже клеточные формы жизни — эукариот и прокариот.
Как известно, в состав клеток входят ДНК и РНК, в то время как вирусы содержат только один тип нуклеиновой кислоты — ДНК или РНК. Поэтому все вирусы подразделяют на две группы — ДНК-геномные и РНК-геномные. Обычно вирусы растений содержат РНК-геномы, вирусы человека и животных как ДНК-, так и РНК-геномы. Почти все бактериофаги ДНК-геномны.
Сложно организованные вирусы (вирусы животных и человека) сложны по химическому составу и содержат дополнительные белковые или липопротеидные оболочки. Кроме нуклеиновой кислоты и белков, они содержат липиды в наружных оболочках и углеводы в составе белков наружных оболочек (гликопротеидов). Некоторое количество липидов есть у бактериофагов и ряда крупных вирусов растений. У некоторых сложных вирусов выявлены ферменты. У бактериофагов также обнаружены ферменты — лизоцим и аденозинтрифосфатаза.
Один из наиболее хорошо изученных фитопатогенных вирусов — вирус табачной мозаики (ВТМ). В 1935 г. У. Стенли выделил и получил этот вирус в кристаллической форме. При введении в рас-

Рис. 28. Электронные микрофотографии вирусов животных, растений и бактерий: слева — коровьей оспы, заболеваний насекомых, бактериофага ТЗ. полиомиелита; справа — гриппа, бактериофага Г2. папилломы кроликов, мозаики табака
тение табака кристаллы вызывали симптомы мозаичной болезни. Получены в кристаллическом виде и многие другие вирусы.
Изучение вирусов под электронным микроскопом показало, что они разнообразны по форме и имеют довольно сложное строение. Различают следующие формы вирусов: палочковидную, при которой вирус имеет вид прямого цилиндра (вирус табачной мозаики); нитевидную, представляющую эластичные изгибающиеся нити (некоторые вирусы растений и бактерий); сферическую, сходную с многогранниками (вирусы животных и человека); кубовидную, по виду напоминающую параллелепипед с закругленными краями (вирусы животных и человека); булавовидную, характеризующуюся наличием головки и отростка (вирусы бактерий и актиномицетов) (рис. 28).
Внеклеточная форма существования вируса, вирион, состоит из нуклеиновой кислоты и белка. Нуклеиновая кислота уложена в виде спирали и окружена белковой оболочкой, называемой капсидом. Последний образован большим числом субъединиц белка — капсамеров, которые, в свою очередь, представлены одной или несколькими молекулами белка. Белковый капсид, объединенный с нуклеиновой кислотой (ДНК или РНК), носит название нуклеокапсида. По способу укладки капсомсров выделяют капсиды, построенные по спиральному и кубическому типам симметрии. В первом случае капсид имеет цилиндрическую форму, во втором — форму многогранника. К вирусам со спиральным типом симметрии относят вирус табачной мозаики.

Рис. 29. Т-бактериофаг. Электронная микрофотография (по: С. Бреннер)
Для многих вирусов бактерий, или фагов, характерен так называемый сложный тип симметрии: головка фага имеет форму многогранника (кубическая симметрия), хвостовой отросток — форму цилиндра (спиральная симметрия) (рис. 29).
Размеры вирусов определяют различными способами: по размеру пор фильтров, пропускающих вирусы, по скорости осаждения вирусов при центрифугировании и при помощи фотографий, полученных в электронном микроскопе. Размеры вирионов вирусов колеблются в довольно широких пределах — от 15 до 400 нм. В обычный световой микроскоп отдельные вирусные частицы не видны, но в пораженных вирусом клетках часто можно различить тельца-включения, представляющие собой, как считают, гигантские колонии вирусов.
Вирусы специфичны, они паразитируют только на определенных хозяевах — растениях, животных или микроорганизмах. Это обусловливает распределение вирусов на группы на основе типа хозяев. В последнее время при классификации вирусов принимают во внимание их строение, чувствительность к внешним факторам и т. д. Выделяют группы вирусов, патогенных для растений, животных и, наконец, для микроорганизмов. Вирусы бактерий и актиномицетов называют соответственно бактериофагами и актииофагами. Известны субмикроскопические агенты — микофаги, поражающие грибы, и цианофаги, паразитирующие на цианобактериях.
Вирусы нс размножаются в почве, но могут долго сохраняться в ней, если условия исключают их инактивацию. Так сохраняются вирусы мозаичной болезни пшеницы, овса и табака, кольцевой пятнистости картофеля и др. Некоторые вирусы человека и животных, попадая в почву, остаются инфекционными в течение нескольких месяцев.
Фаги — облигатные паразиты микроорганизмов — открыли независимо друг от друга в 1915 г. Ф. Туорт ив 1917 г. Ф. Д. Эррель. Длина головки фага достигает 60—100 нм, отростка — 100—200 нм. Призматическая головка фага покрыта оболочкой из упорядоченно расположенных капсомсров. Внутри головки находится одна или две нити ДНК.
Отросток представляет собой белковый стержень, покрытый сверху чехлом из спирально расположенных капсомеров, способных к сокращению. Обычно отросток оканчивается базальной пластинкой с пятью-шестью выростами. От пластинки отходят тонкие нити — органы адсорбции. Через отросток из головки фага ДНК переходит в клетку микроорганизма.
Механизм проникновения бактериофага в бактерии подробно изучен. Обычно фаг адсорбируется чувствительной к нему клеткой бактерии. Затем содержимое головки (ДНК) переходит в бактерию, а оболочка остается снаружи. После нападения фага бактерия утрачивает способность к делению, перестает двигаться. Метаболизм бактериальной клетки перестраивается под влиянием ДНК фага, и клетка начинает производить продукты не собственного обмена, а бактериофага, и в результате в ней происходит интенсивное образование частиц бактериофага. Затем клеточная стенка бактерии растворяется, и из нес выходят зрелые бактериофаги. Одна клетка бактерии становится источником нескольких сотен и даже тысяч бактериофагов.
При наблюдении колоний бактерий на агаре лизирующее действие бактериофага видно по образованию прозрачных зон вокруг колоний, а на жидкой среде — но уменьшению мутности бактериальной суспензии.
Растворять (лизировать) данный вид бактерий способен только вирулентный к нему фаг. Нередко бактериальная клетка инфицируется фагом, который может в ней существовать, не вызывая лизиса. При размножении бактерии инфекционное начало переходит в дочерние клетки. Бактериофаги такого характера называют умеренными, а бактерий — передатчиков данных фагов — лизогенными. При определенных условиях лизогенные культуры бактерий могут быть лизированы находящимся в них фагом. Каждый фаг способен поражать бактерий одного вида или группы близких видов.
Исследовано большое число фагов, поражающих рахзичных микроорганизмов. Известны фаги, лизирующие бактерии родов Pseudomonas, Bacillus, Rhizobium, Streptococcus, Staphylococcus; актиноми исты рода Streptomyces; микобактерии рода Mycobacterium и др. Фаги встречаются в воде, почве и других природных объектах. Некоторых фагов используют в медицине для профилактики заболеваний.
1. Назовите основные группы водорослей и их свойства. 2. Какие группы простейших широко представлены в почве? 3. Чем отличаются микромицс- ты от миксомицетов? 4. Что представляют собой вирусы и какие организмы они способны заражать?
Морфология вирусов
Вирусы являются самыми мелкими из всех микроорганизмов. Для них принято измерение в миллимикронах и в ангстремах. Для определения таких размеров частиц применяется несколько методов. Так, взвесь вирусов пропускают через особые фильтры из коллодия, имеющие очень мелкие поры определенной величины. Фильтрование производят через несколько фильтров с разной величиной пор. Разница между диаметрами пор последнего фильтра, пропустившего вирусные частицы, и фильтра, уже не пропустившего вирусные частицы, указывает средние размеры вирусных частиц. При сверхскоростном центрифугировании (50 и более тысяч оборотов в минуту) величина вирусных частиц определяется по специальной формуле в зависимости от числа оборотов и времени осаждения частиц. При этом происходит и очистка вируса от посторонних веществ. Для этого подбирают такие скорости, при которых выпадают посторонние частицы, сначала крупные, а потом самые мелкие. При наиболее высокой скорости получают лишь частицы вируса.
Человек увидел вирусы только после 1940 г., когда был построен и усовершенствован электронный микроскоп. При увеличении в десятки и сотни тысяч раз удалось изучить форму, величину, а также строение частиц некоторых вирусов.
Было найдено, что как величина, так и форма отдельных особей (элементарных частиц) разных видов вирусов довольно разнообразны. Имеются крупные вирусы (например, вирус орнитоза, оспы, трахомы и др.), вирусы средних размеров (гриппа, чумы, бешенства) и мелкие (вирус полиомиелита, кори, ящура, энцефалитов, вирусы многих растений). В таблице приводятся размеры некоторых вирусов, определенные разными способами, в миллимикронах (по В. М. Жданову и Шен).
Самые крупные вирусы приближаются по своим размерам к самым мелким бактериям, а самые мелкие вирусы близки к крупным белковым молекулам.
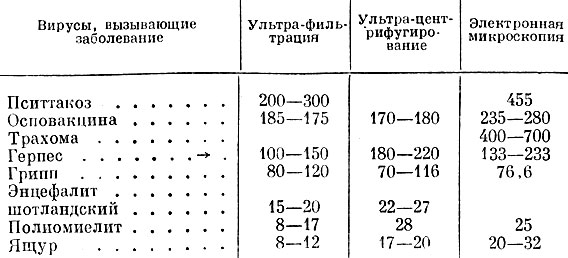
Таблица 7
По внешнему виду одни вирусы имеют шарообразную форму (вирус гриппа), другие - кубовидную форму (вирус оспы), третьи - вид палочки. Вирус табачной мозаики (ВТМ) имеет вид тонкой шестигранной палочки длиной 300 ммк и в диаметре 15 ммк.

Рис. 42. Сравнительная величина бактерий, риккетсий, вирусов и белковых молекул (из Риверса, 1952, с изменениями)
При многих вирусных инфекциях (оспа, бешенство, трахома и др.) наблюдаются в цитоплазме или ядре клетки организма хозяина особые, специфические для каждой инфекции внутриклеточные тельца - включения. Они довольно крупные, и их можно видеть в световой микроскоп.
В большинстве случаев включения представляют собой скопление элементарных телец, вирусных частиц, как бы их колонию. Наличие их в клетках помогает при диагностике некоторых заболеваний.
Одним из своеобразных свойств многих вирусов растений является их способность образовывать кристаллы. Д. И. Ивановский первый наблюдал в листьях табака, пораженных ВТМ, включения, называемые теперь кристаллами Ивановского. Они состоят из элементарных частиц вируса табачной мозаики. Кристаллы вируса можно растворять, как растворяют сахар, соль. Из раствора этот вирус можно выделить в аморфном, некристаллическом, состоянии. Осадок можно вновь растворить, затем снова превратить в кристаллы. Если растворить кристаллический вирус в тысячу раз, то капля такого раствора вызовет у растения мозаичную болезнь. Из вирусов человека и животных пока получены кристаллы вируса полиомиелита. Каждый кристалл состоит из миллионов вирусных частиц.
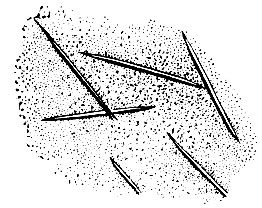
Рис. 43. Кристаллы ВТМ
Химический состав вирусов был изучен прежде всего у возбудителя табачной мозаики. Этот вирус представляет собой чистый нуклеопротеид, т. е. состоит из белка и нуклеиновой кислоты. Вирусный нуклеопротеид табачной мозаики имеет огромный молекулярный вес (40-50 млн.).
Вирусная частица имеет сложное строение. Нуклеиновая кислота находится внутри вирусной частицы, она окружена белковой оболочкой. В вирусной частице обычно содержится одна молекула нуклеиновой кислоты.
Вирусы растений содержат рибонуклеиновую кислоту, фаги содержат дезоксирибонуклеиновую кислоту. В вирусах человека и животных находится или РНК, или ДНК. РНК содержится в вирусах гриппа (1,6%), полиомиелита (24%), некроза табака (18%), мозаики табака (6%), ящура (40%), саркомы Рауса (10%) и др. ДНК содержится в вирусах осповакцин (6%), папилломы (6,8%), герпеса (3,8%), полиомы (12%) и др.
Теперь интенсивно изучается вопрос, как соединяются белок и нуклеиновая кислота, как они подогнаны друг к другу. Для разрешения этого вопроса пользуются методом рентгенокристаллографии. Если в вирусной частице имеются субъединицы, то этот метод может установить их число, а также их взаимное расположение. Оказалось, что для большинства вирусов характерно закономерное, высокоупорядоченное расположение элементов вирусной частицы.
У вируса полиомиелита нуклеиновая кислота свернута в клубок, белковая оболочка состоит из 60 одинаковых субъединиц, которые объединены в 12 групп, по 5 субъединиц в каждой. Частица вируса имеет сферическую форму.
Нуклеиновая кислота вируса табачной мозаики имеет вид спирали или пружины. Белковая оболочка ВТМ состоит также из отдельных одинаковых по форме и размерам белковых субъединиц. Всего имеется 2200 субъединиц, расположенных в виде 130 витков вокруг стержня нуклеиновой кислоты. Молекулярный вес такой субъединицы 18 000. Каждая субъединица представляет собой пептидную цепочку, содержащую 158 определенных аминокислот, причем уже определено последовательное расположение этих аминокислот. В настоящее время интенсивно изучается последовательность расположения 6500 нуклеотидов, образующих нуклеиновую кислоту. Когда эта задача будет решена, то станет известен план, которым определяется тип вируса, образующегося в зараженной клетке. Строение, подобное частицам ВТМ и полиомиелита, имеют другие мелкие вирусы растений.
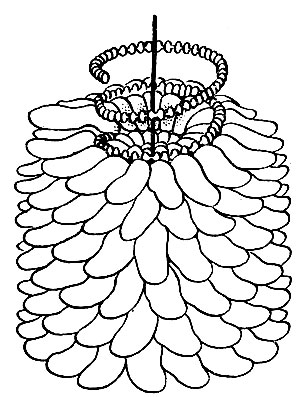
Рис. 44. Схема строения частицы ВТМ, основанная на данных рентгеноструктурного анализа, проведенного Франклином. Для ясности части цепи рибонуклеиновой кислоты показаны без опорного белкового каркаса. На каждую белковую единицу приходится 3 нуклеотида, или 49 на оборот спирали. Каждый нуклеотид схематически представлен плоским диском, поскольку в этом направлении ориентированы пуриновые и пиримидиновые основания (Франклин, 1955)
У более крупных вирусов, кроме нуклеиновой кислоты, белковой оболочки, есть еще внешние оболочки, содержащие белки, липоиды, углеводы. Некоторые вирусы содержат ферменты. Так, гриппозный вирус имеет фермент нейраминидазу, парагриппозный вирус - сендай-лизин, вирус миелобластоза птиц содержит аденовинтрифосфатазу. Эти ферменты растворяют оболочку клеток для проникновения вируса в тело своего будущего хозяина.
Другой особенностью вирусов является их внутриклеточный паразитизм. Начиная с Ивановского никому из ученых не удавалось выращивать вирусы на таких искусственных питательных средах, на которых так пышно размножаются самые требовательные паразиты из бактерий. Вирусы развиваются и размножаются только внутри живых клеток поражаемых ими организмов, которые называются хозяевами. Разные вирусы приспособились к паразитированию у определенных хозяев. Так, вирус полиомиелита размножается только в организме человека и мыши; вирус кори поражает только людей. Кроме того, размножение вируса происходит не во всех тканях организма хозяина и неодинаково в разных тканях: гриппозный вирус хорошо развивается в легочной ткани; вирус бешенства - нервной ткани; вирус трахомы обитает только на слизистой оболочке глаза. Таким образом, вирусы обладают специфичностью. Но многие вирусы растений могут паразитировать на разных видах растений. Так, ВТМ (вирус табачной мозаики) поражает почти все виды пасленовых растений. Многие вирусы растений хорошо размножаются в тканях насекомых, которые переносят вирус с одного растения на другое.

Рис. 45. Вирус табачной мозаики (увеличено в 230 000 раз)
В свободном состоянии, во внешней среде вне живой клетки, вирусы не проявляют активности, они только сохраняют свою жизнеспособность, иногда продолжительное время. Но как только вирусы встречаются с чувствительными к ним клетками, они становятся активными, внедряются в них и проявляют все признаки жизнедеятельности.
Единственным методом изучения жизнедеятельности вирусов раньше было заражение восприимчивых к ним опытных животных: мышей, кроликов, обезьян и др. Более удобно и экономно выращивать вирусы в развивающемся зародыше куриного яйца. Материал, содержащий вирус, вводят шприцем в зародыш на 8-12-й день его развития. Через несколько дней пребывания зародыша в термостате изучают патологические изменения, вызываемые вирусом в зародыше. Затем пересевают в свежий зародыш другого яйца. В последнее время получил наибольшее применение метод однослойных культур из изолированных клеток животных тканей. Размельченную свежую ткань обрабатывают ферментом трипсином, который разрушает межклеточные связи. Освободившиеся клетки отмывают от трипсина, разводят питательным составом (№ 199, содержащим необходимые аминокислоты и соли) и помещают в пробирки или в специальные плоские чашки. В термостате клетки размножаются, образуя однослойный пласт по стеклу. Затем эту культуру однородных клеток заражают вирусом и происходящие в ней процессы изучают под микроскопом или другими способами. Так трудоемкий и дорогой способ, например культура вируса полиомиелита на печени обезьян, был заменен быстрым способом выращивания его в тканевой культуре.
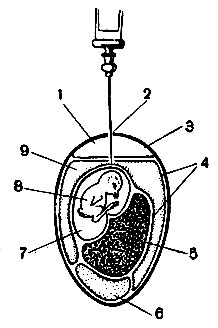
Рис. 46. Метод заражения куриного эмбриона. Заражение в аллантоисную полость куриного эмбриона: 1 - воздушное пространство; 2 - отверстия в скорлупе; 3 - скорлупная оболочка; 4 - хорион-аллантоисная оболочка; 5 - желточный мешок; 6 - белок; 7 - амнион; 8 - плод; 9 - аллантоисная полость
В 1955 г. и позднее были получены необычные факты, вызвавшие недоумение у ученых биологов. Химическим путем вирус табачной мозаики был разделен на свои составные части: белок и нуклеиновую кислоту. Каждая из них в отдельности не вызывала заболевания мозаикой у листьев табака. Но когда их опять соединили вместе в пробирке (10 частей белка и 1 часть нуклеиновой кислоты) и заразили этой смесью листья табака, то получили типичную мозаику на листьях, как от исходного цельного ВТМ. При электронной микроскопии были найдены типичные палочки вируса, состоящие из белковой оболочки, в которой был заключен тяж нуклеиновой кислоты. Таким образом, нуклеиновая кислота связалась с белковой частью и заняла в ней свое нормальное положение. Открытие этого явления - ресиитеза (восстановления) вируса - является крупнейшим достижением современной микробиологии, открывающим новые пути в биологии и медицине.
Далее оказалось, что достаточно натереть лист табака лишь одной нуклеиновой кислотой, выделенной из ВТМ мягким способом, как на листе возникают (конечно, не в большом количестве) типичные некрозы, в которых находились в огромном количестве типичные цельные вирусные частицы.
Такие же результаты были получены с вирусами человека: полиомиелита, гриппа и др.
Выл получен даже гибридный вирус мозаики табака из белка одного типа вируса и РНК другого типа вируса, отличавшегося по некоторым признакам от вируса первого типа. При размножении этот гибридный вирус давал потомство только того вируса, чью РНК содержал гибрид.
Все эти факты говорят о том, что нуклеиновым кислотам принадлежит ведущая роль в размножении вирусов и инфекциозности их. Нуклеиновые кислоты обеспечивают передачу наследственных свойств. В кислотах заключена наследственная информация по синтезу полноценных вирусных частиц внутри клетки.
Белковая оболочка вируса несет защитную функцию, охраняя от внешних воздействий хрупкую нить нуклеиновой кислоты, кроме того, помогает вирусу проникать в клетку, определяет специфичность вирусов. Но некоторые ученые не считают возможным так ограничивать значение белков. Нужны дальнейшие исследования о роли вирусных белков.
Процесс размножения вирусов принципиально отличается от процесса размножения бактерий, простейших и других клеточных организмов.
Различают четыре фазы этого процесса: прикрепление вирусных частиц к клетке хозяина, проникновение вируса внутрь клетки, внутриклеточное размножение вируса и выход новых частиц вируса из клетки.
Первая фаза - прикрепление, или адсорбция, вируса к клетке - изучена в отношении вирусов гриппа и полиомиелита. Стенка клетки имеет мозаичное строение, в одних местах ее выступают молекулы мукопротеидов, в других молекулы липопротеидов. Вирус гриппа адсорбируется на мукопротеидах, а вирус полиомиелита адсорбируется на липопротеидах. Адсорбцию можно наблюдать в электронный микроскоп. В месте адсорбции вируса на стенке клетки образуется углубление, куда втягивается частица вируса. Края углубления смыкаются, и частица вируса оказывается внутри клетки (виропексис). Одновременно с виропексисом происходит разрушение белковой оболочки вируса. Проникновению вируса гриппа в клетку способствует фермент его оболочки. Таким образом, в клетку проникает нуклеиновая кислота, освобожденная от белковых оболочек с помощью ферментов самой клетки.
В третьей фазе проникшая в клетку вирусная нуклеиновая кислота включается в обмен веществ клетки и направляет аппарат синтеза клетки на производство белка и нуклеиновой кислоты не клетки, а новых вирусных частиц. Деятельность ферментов, участвующих в синтезе вируса, активизируется, а остальных ферментов тормозится. Кроме того, создаются новые ферменты, которых клетка не имела, но которые необходимы для синтеза вирусных частиц. Можно полагать, что в это время организуется новая единая система вирус - клетка, переключенная на синтез вирусного материала. В начале этой фазы не удается различить в клетке какие-либо элементы вируса.
Обычно нуклеиновые кислоты и белки вируса синтезируются не одновременно и в разных местах клетки. Сначала начинается синтез нуклеиновой кислоты, а затем несколько позднее идет синтез белка. После накопления этих составных частей вируса происходит их объединение, сборка в полноценные вирусные частицы. Иногда образуются неполные вирусные частицы, лишенные нуклеиновой кислоты и потому неспособные к самопроизводству (бублики).
Быстро наступает последняя фаза - выход вирусных частиц из клетки. В каком-либо месте клетки сразу выходит около 100 частиц вируса, У более сложных вирусов имеются еще внешние оболочки вирусного нуклеопротеида, которыми они обволакиваются во время прохождения через клетку и выхода из нее, при этом в состав внешних оболочек входят белки клетки хозяина.
У вирусов человека и животных выход нового потомства происходит в несколько циклов. Так, у вируса гриппа каждый цикл продолжается часов 5-6 с выходом 100 и более вирусных частиц одной клетки, а всего наблюдается 5-6 циклов в течение 30 часов. После этого способность клетки производить вирус истощается, и она погибает. Весь процесс размножения парагриппозного вируса Сен дай от адсорбции до выхода из клетки продолжается 5-6 часов.
Иногда частицы вируса не выходят из клетки, а скапливаются в ней в виде внутриклеточных включений, очень характерных для разных видов вирусов. Вирусы растений образуют включения, имеющие кристаллическую форму.
Большое внимание начинает привлекать к себе семейство микробов, получившее название "микоплазма", так как за последнее время в этой группе обнаружены возбудители различных заболеваний человека и животных. В виде скрытой инфекции они часто обитают во многих тканевых культурах - Хела и др. Микоплазмы занимают промежуточное положение между бактериями и вирусами. С вирусами их сближает фильтруемость через бактериальные фильтры, фильтрующиеся формы способны к саморепродукции, внутриклеточному размножению. К признакам, сближающим вирусы с бактериями, относится способность расти на питательных средах, образовывать на них колонии, а также отношение к антибиотикам, сульфамидам и их антигенная структура.
Читайте также:
