
Основные схемы репликации вирусов при продуктивной инфекции
Вирусы — облигатные внутриклеточные паразиты, способные только к внутриклеточному размножению. В вирусинфицированной клетке возможно пребывание вирусов в различных состояниях:
воспроизводство многочисленных новых вирионов;
пребывание нуклеиновой кислоты вируса в интегрированном состоянии с хромосомой клетки (в виде провируса);
существование в цитоплазме клетки в виде кольцевых нуклеиновых кислот, напоминающих плазмиды бактерий.
Поэтому диапазон нарушений, вызываемых вирусом, весьма широк: от выраженной продуктивной инфекции, завершающейся гибелью клетки, до продолжительного взаимодействия вируса с клеткой в виде латентной инфекции или злокачественной трансформации клетки.
Различают три типа взаимодействия вируса с клеткой: продуктивный, абортивный и интегративный.
1. Продуктивный тип — завершается образованием нового поколения вирионов и гибелью (лизисом) зараженных клеток (цитоли-тическая форма). Некоторые вирусы выходят из клеток, не разрушая их (нецитолитическая форма).
Абортивный тип — не завершается образованием новых вирионов, поскольку инфекционный процесс в клетке прерывается на одном из этапов.
Интегративный тип, или вирогения —характеризуется встраиванием (интеграцией) вирусной ДНК в виде провируса в хромосому клетки и их совместным сосуществованием (совместная репликация).
Репродукция вирусов (продуктивный)
1) адсорбция вирионов на клетке;
2) проникновение вируса в клетку;
4) синтез вирусных компонентов;
5) формирование вирионов;
6) выход вирионов из клетки.
У различных вирусов эти стадии отличаются
Специфические рецепторы клеток имеют различную природу, являясь белками, липидами, углеводными компонентами белков, липидов и др. Так, рецепторами для вируса гриппа является сиаловая кислота в составе гли-копротеинов и гликолипидов (ганглиозидов) клеток дыхательных путей. Вирусы бешенства адсорбируются на ацетилхолиновых рецепторах нервной ткани, а вирусы иммунодефицита человека — на СО4-рецепторах Т-хелперов, моноцитов и дендритных клеток. На одной клетке находится от десяти до ста тысяч специфических рецепторов, поэтому на ней могут адсорбироваться десятки и сотни вирионов.
Наличие специфических рецепторов лежит в основе избирательности поражения вирусами определенных клеток, тканей и органов. Это так называемый тропизм (греч. tropos — поворот, направление). Например, вирусы, репродуцирующиеся преимущественно в клетках печени, называются гепатотропными, в нервных клетках — нейротропными, в иммунокомпетентных клетках — иммунотропными и т. д.
Проникновение вирусов в клетку. Вирусы проникают в клетку путем рецептор-зависимого эндоцитоза (виропексиса), или слияния оболочки вируса с клеточной мембраной, или же в результате сочетания этих механизмов.
2. Слияние обточки вириона с клеточной мембраной характерно только для некоторых оболочечных вирусов (парамиксовирусов, ретровиру-сов, герпесвирусов), в составе которых имеются белки слияния. Происходит точечное взаимодействие вирусного белка слияния с липидами клеточной мембраны, в результате чего вирусная липопротеиновая оболочка интегрирует с клеточной мембраной, а внутренний компонент вируса попадает в цитозоль.
В) Синтез вирусных компонентов. Синтез белков и нуклеиновых кислот вируса, который разобщен во времени и пространстве. Синтез осуществляется в разных частях клетки, поэтому такой способ размножения вирусов называется дизъюнктивным (от лат. disjunctus — разобщенный).
С)Синтез вирусных белков. В зараженной клетке вирусный геном кодирует синтез двух групп белков:
1. неструктурных белков, обслуживающих внутриклеточную репродукцию вируса на разных его этапах;
2. структурных белков, которые входят в состав вириона (геномные, связанные с геномом вируса, капсидные и су-перкапсидные белки).
К неструктурным белкам относятся: 1) ферменты синтеза РНК или ДНК (РНК- или ДНК-полимеразы), обеспечивающие транскрипцию и репликацию вирусного генома; 2) белки-регуляторы; 3) предшественники вирусных белков, отличающиеся своей нестабильностью в результате быстрого нарезания на структурные белки; 4) ферменты, модифицирующие вирусные белки, например, протеиназы и протеинкиназы.
I. ДНК-содержашие вирусы реализуют генетическую информацию так же, как и клеточный геном, по схеме:
Причем ДНК-содержашие вирусы используют для этого процесса клеточную полимеразу (вирусы, геномы которых транскрибируются в ядре клетки — аденовирусы, па-повавирусы, герпесвирусы) или собственную РНК-полимеразу (вирусы, геномы которых транскрибируются в цитоплазме, например поксвирусы).
II. Плюс-нитевые РНК-содержашие вирусы (например, пикорнавирусы, флавивирусы, тогавирусы) имеют геном, выполняющий функцию иРНК; он распознается и транслируется рибосомами. Синтез белков у этих вирусов осуществляется без акта транскрипции по схеме:
геномная РНК вируса -> трансляция белка вируса.
III. Геном минус-однонитевых РНК-содержаших вирусов (ортомиксовирусов, парамиксовирусов, рабдовирусов) и двунитевых (реовирусов) служит матрицей, с которой транскрибируется иРНК, при участии РНК-полимеразы, связанной с нуклеиновой кислотой вируса. Синтез белка у них происходит по схеме:
IV. Ретровирусы (вирусы иммунодефицита человека, онкогенные ретровирусы) имеют уникальный путь передачи генетической информации. Геном ретровирусов состоит из двух идентичных молекул РНК, т. е. является диплоидным. В составе ретровирусов есть особый вирусоспецифический фермент — обратная транскриптаза, или ревертаза, с помощью которой осуществляется процесс обратной транскрипции, т. е. на матрице геномной РНК синтезируется комплементарная однонитевая ДНК (кДНК). Комплементарная нить ДНК копируется с образованием двунитевой комплементарной ДНК, которая интегрирует в клеточный геном и в его составе транскрибируется в иРНК с помощью клеточной ДНК-зависимой РНК-полимеразы. Синтез белков для этих вирусов осуществляется по схеме:
Репликация вирусных геномов, т. е. синтез вирусных нуклеиновых кислот, приводит к накоплению в клетке копий исходных вирусных геномов, которые используются при сборке вирионов. Способ репликации генома зависит от типа нуклеиновой кислоты вируса, наличия вирусоспецифических или клеточных полимераз, а также от способности вирусов индуцировать образование полимераз в клетке.
Механизм репликации отличается у вирусов, имеющих:
1) двунитевую ДНК;
2) однонитевую ДНК;
3) плюс-однонитевую РНК;
4) минус-одноните-вую РНК;
5) двунитевую РНК;
6) идентичные плюс-нитевые РНК (ретровирусы).
1. Двунитевые ЛНК-вирусы. Репликация двунитевых вирусных ДНК происходит обычным полуконсервативным механизмом: после рас- плетения нитей ДНК к ним комплементарно достраиваются новые нити. Каждая вновь синтезированная молекула ДНК состоит из одной родительской и одной вновь синтезированной нити. К этим вирусам относится большая группа вирусов, которые содержат двунитевую ДНК в линейной (например, герпесвирусы, аденовирусы и поксвирусы) или в кольцевой форме, как папилломавирусы. У всех вирусов, кроме поксвирусов, транскрипция вирусного генома происходит в ядре.
Уникальный механизм репликации характерен для гепаднавирусов (вируса гепатита В). Геном гепаднавирусов представлен дву-нитевой кольцевой ДНК, одна нить которой короче (неполная плюс-нить) другой нити. Первоначально достраивается (рис. 3.7). Затем полная двунитевая ДНК с помощью клеточной ДНК-зависимой РНК-полимеразы транскрибируется с образованием небольших молекул иРНК и полной однонитевой плюс-РНК. Последняя называется прегеномной РНК; она является матрицей для репликации генома вируса. Синтезированные иРНК участвуют в процессе трансляции белков, в том числе вирусной РНК-зависимой ДНК-полимеразы (обратной транскриптазы). С помощью этого фермента мигрирующая в цитоплазму прегеномная РНК обратно транскрибируется в минус-нить ДНК, которая, в свою очередь, служит матрицей для синтеза плюс-нити ДНК. Этот процесс заканчивается образованием двунитевой ДНК, содержащей неполную плюс-нить ДНК.
Однонитевые ДНК-вирусы. Единственными представителями однонитевых ДНК-вирусов являются парвовирусы. Парвовирусы используют клеточные ДНК-полимеразы для создания двунитевого вирусного генома, так называемой репликативной формы послед него. При этом на исходной вирусной ДНК (плюс-нить) комплементарно синтезируется минус-нить ДНК, служащая матрицей для синтеза плюс-нити ДНК нового вириона. Параллельно синтезируется иРНК, происходит трансляция вирусных пептидов.
Плюс-однонитевые РНК-вирусы. Эти вирусы включают большую группу вирусов — пикорнавирусы, флавивирусы, тогавирусы (рис.3.8), у которых геномная плюс-нить РНК выполняет функцию иРНК. Например, РНК полиовирусов после проникновения в клетку связывается с рибосомами, работая как иРНК, и на ее основе синтезируется большой полипептид, который расщепляется на фрагменты: РНК-зависимую РНК-полимеразу, вирусные протеазы и капсидные белки. Полимераза на основе геномной плюс-нити РНК синтезирует минус-нить РНК; формируется временно двойная РНК, названная промежуточным репликативным звеном. Это промежуточное репликативное звено состоит из полной плюс-нити РНК и многочисленных частично завершенных минус-нитей. Когда образованы все минус-нити, они используются как шаблоны для синтеза новых плюс-нитей РНК. Этот механизм используется как для размножения геномной РНК вируса, так и для синтеза большого количества вирусных белков.
Минус-однонитевые РНК-вирусы. Минус -однонитевые РНК-вирусы (рабдовирусы, парамиксовирусы, ортомиксовирусы) имеют в своем составе РНК-зависимую РНК-полимеразу. Проникшая в клетку геномная минус- нить РНК трансформируется вирусной РНК-зависимой РНК-полимеразой в неполные и полные плюс-нити РНК. Неполные копии выполняют роль иРНК для синтеза вирусных белков. Полные копии являются матрицей (промежуточная стадия) для синтеза минус-нитей геномной РНК потомства
Двунитевые РНК-вирусы. Механизм репликации этих вирусов (реовирусов и ротави-русов) сходен с репликацией минус-однонитевых РНК-вирусов. Отличие состоит в том, что образовавшиеся в процессе транскрипции плюс-нити функционируют не только как иРНК, но и участвуют в репликации: они являются матрицами для синтеза минус-нитей РНК. Последние в комплексе с плюс-нитями РНК образуют геномные двунитевые РНК вирионов. Репликация вирусных нуклеиновых кислот этих вирусов происходит в цитоплазме клеток.
6. Ретровирусы (плюс-нитевые диплоидные РНК-содержащие вирусы). Обратная транс-криптаза ретровирусов синтезирует (на матрице РНК-вируса) минус-нить ДНК, с которой копируется плюс-нить ДНК с образованием двойной нити ДНК, замкнутой в кольцо (рис. 3.10). Далее двойная нить ДНК интегрирует с хромосомой клетки, образуя провирус. Многочисленные вирионные РНК образуются в результате транскрипции одной из нитей интегрированной ДНК при участии клеточной ДНК-зависимой РНК-полимеразы.
Формирование вирусов. Вирионы формируются путем самосборки: составные части вириона транспортируются в места сборки вируса — участки ядра или цитоплазмы клетки. Соединение компонентов вириона обусловлено наличием гидрофобных, ионных, водородных связей и стерического соответствия.
Существуют следующие общие принципы сборки вирусов:
Формирование вирусов — многоступенчатый процесс с образованием промежуточных форм, отличающихся от зрелых вирионов по составу полипептидов.
Сборка просто устроенных вирусов заключается во взаимодействии вирусных нуклеиновых кислот с капсидными белками и в образовании нуклеокапсидов.
У сложноустроенных вирусов сначала формируются нуклеокапсиды, которые взаимодействуют с модифицированными мембранами клеток (будущей липопротеиновой оболочкой вируса).
Причем сборка вирусов, реплицирующихся в ядре клетки, происходит с участием мембраны ядра, а сборка вирусов, репликация которых идет в цитоплазме, осуществляется с участием мембран эндоплазматической сети или плазматической мембраны, куда встраиваются гликопротеины и другие белки оболочки вируса.
У ряда сложноустроенных вирусов минус-нитевых РНК-вирусов (ортомиксовирусов, парамиксовирусов) в сборку вовлекается так называемый матриксный белок (М-белок), который расположен под модифицированной клеточной ембраной. Обладая гидрофобными свойствами, он выполняет роль посредника между нуклеокапсидом и вирусной липопротеиновой оболочкой.
□ Сложноустроенные вирусы в процессе формирования включают в свой состав некоторые компоненты клетки хозяина, например липиды и углеводы.
Выход вирусов из клетки. Полный цикл репродукции вирусов завершается через 5—6 ч (вирус гриппа и др.) или через несколько суток (гепатовирусы, вирус кори и др.). Процесс репродукции вирусов заканчивается выходом их из клетки, который происходит взрывным путем или почкованием, экзоцитозом.
Взрывной путь: из погибающей клетки одновременно выходит большое количество вирионов. По взрывному пути выходят из клетки просто устроенные вирусы, не имеющие липопротеиновой оболочки.
Почкование, экзоцшпт присущи вирусам, имеющим липопротеиновую оболочку, которая является производной от клеточных мембран. Сначала образовавшийся нуклеокапсид или сердцевина вириона транспортируется к клеточным мембранам, в которые уже встроены вирусоспецифические белки. Затем в области контакта нуклеокапсида или сердцевины вириона с клеточной мембраной начинается выпячивание этих участков. Сформировавшаяся почка отделяется от клетки в виде сложно устроенного вируса. При этом клетка способна длительно сохранять жизнеспособность и продуцировать вирусное потомство.
Почкование вирусов, формирующихся в цитоплазме, может происходить либо через плазматическую мембрану (например, парамиксовирусы, тогавирусы), либо через мембраны эндоплазматической сети с последующим их выходом на поверхность клетки (например, буньявирусы).
Вирусы, формирующиеся в ядре клетки (например, герпесвирусы), почкуются в перинуклеарное пространство через модифицированную ядерную мембрану, приобретая таким образом липопротеиновую оболочку. Затем они транспортируются в составе цитоплазма-тических везикул на поверхность клетки.
Взаимодействие вируса с клеткой хозяина - это сложный «многоступенчатый процесс, который начинается с адсорбции вирусных частиц на рецепторах клетки хозяина и продолжается после их проникновения внутрь клетки. В результате такого взаимодействия мовивается либо продуктивная, либо абортивная, либо интегративная форма клеточной инфекции. При продуктивной форме происходит размножение, точнее, репродукция (лат. reproduce - воспроизводить) вируса, при абортивной - ее нарушение на одном из этапов, при интегративной - интеграция вирусной нуклеиновой кислоты в клеточный геном.
Продуктивная инфекция. Репродукция вирусов.
В 50-х годах было установлено, что размножение, или репродукция, вирусов происходит путем репликации их нуклеиновой кислоты и биосинтеза белков с последующей самосборкой вириона. Этот процесс происходит в разных частях клетки - ядре или цитоплазме, вследствие чего получил название дизъюнктивного, т.е. разобщенного размножения.
Вирусная репродукция представляет собой уникальную форму выражения чужеродной (вирусной) информации в клетках человека и животных, насекомых, растений и бактерий, которая состоит в подчинении клеточных матрично-генетических механизмов вирусной информации.
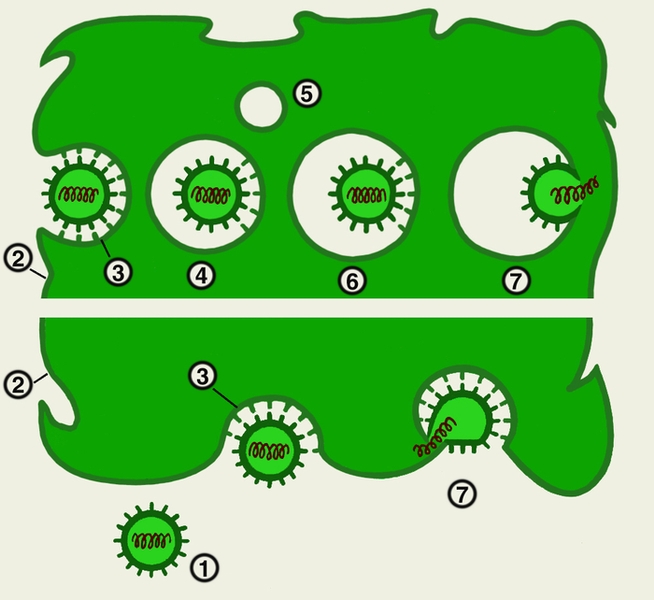
Схема проникновения вирусов в клетку путем рецепторного эндоцитоза (вверху) и через плазматическую мембрану клетки (внизу):
1 — вирусная частица;
2 — плазматическая мембрана клетки;
3 — ямка на поверхности клетки, содержащая рецепторы;
4 — образующаяся клеточная вакуоль;
5 — сливающаяся клеточная вакуоль;
6 — клеточная вакуоль, образующаяся после слияния (рецептосома);
7 — выход генетического материала вируса в цитоплазму.
1-я стадия. Взаимодействие вируса с клеткой начинается с процесса адсорбции, т. е. прикрепления вирусов к поверхности клетки. Это высокоспецифический процесс. Вирус адсорбируется на определенных участках клеточной мембраны — так называемых рецепторах. Клеточные рецепторы могут иметь разную химическую природу, представляя собой белки, углеводные компоненты белков и липидов, липиды. Число специфических рецепторов на поверхности одной клетки колеблется от 10 4 до 10 5 . Следоват ельно, видоспецифичны — вирусы архибактерий не могут инфицировать клетку E. coli, а многие колифаги не проникают в клетку шигеллы. Наиболее выражена тканевая специфичность вирусов животных и человека. Так, вирусы гепатитов поражают гепатоциты, вирус Эпштейна-Барр (вызывает инфекционный мононуклеоз) обладает тропностью к B-лимфоцитам, ВИЧ — к T-лимфоцитам, кишечные вирусы — к энтероцитам, кардиотропностью обладают вирусы Коксаки B. Целый ряд вирусов обладает тропностью не к одному, а к нескольким типам клеток. Так полиовирусы тропны к клеткам респираторного тракта, желудочно-кишечного тракта (ЖКТ), центральной нервной системы (ЦНС). Вирус гепатита C (ВГC) лимфотропен и гепатотропен.
Специфическое сродство вирусов к клеткам и тканям определяется двумя механизмами:
— присутствием на клеточной поверхности специфических для вируса рецепторов.
— содержанием в системе активирующих ферментов, необходимых для протеолитического расщепления вирусных поверхностных белков и проявления инфекционной активности вируса.
2-я стадия - проникновение вируса в клетку хозяина - происходит несколькими путями.
Рецепторно-опосредованный эндоцитоз характеризуется образованием в месте взаимодействия вириона с клеточным рецептором окаймленных пузырьков, в формировании которых принимают участие белки-клатрины.
Виропексис. Этим путем в клетку проникают сложноустроенные вирусы. Он заключается в слиянии мембран - вирусного суперкапсида с клеточной или ядерной мембраной. Данный процесс происходит при помощи специального белка слияния - F-белка, который находится в суперкапсиде. В результате виропексиса капсид оказывается в клетке хозяина, а суперкапсид вместе с белком встраивается в ее плазматическую мембрану (вследствие чего клетка приобретает способность сливаться с другими клетками, что приводит к передаче вируса этим клеткам).
Фагоцитоз. Данным путем вирус проникает в фагоцитирующие клетки, что приводит к незавершенному фагоцитозу.
Проникновение вируса в организм хозяина у разных биологических видов решается по-разному.
1. Вирусы растений проникают в организм хозяина по типу раневых инфекций, где распространяются по плазмодесмам, ксилеме и флоэме.
2. Вирусы бактерий — путем введения нуклеиновой кислоты в тело клетки или путем проникновения вириона.
3. Вирусы насекомых попадают в организм хозяина в процессе питания или размножения.
4. Вирусы животных и человека при инфицировании организма хозяина проходят более сложный путь. Одни вирусы (вирус гриппа, ротавирусы) реплицируются и вызывают заболевание в месте проникновения в организм (входные ворота инфекции). Другие вирусы, попав в организм хозяина с использованием того или иного механизма, проходят стадию распространения.
3-я стадия - транспорт вируса внутри клетки. Он происходит с помощью внутриклеточных мембранных пузырьков, в которых вирус переносится на рибосомы, эндоплазматическую сеть или в ядро.
Распространение вируса в организме сопровождается виремией (вирусемией) — циркуляцией вируса в крови, что свидетельствует о генерализации инфекции.
Различают несколько путей распространения вирусов в организме:
1. Нейронный путь (вирусы бешенства, герпеса).
2. Лимфатический путь (реовирусы, полиомавирусы).
3. Гематогенный путь, ассоциированный с клеточными компонентами и плазмой крови (вирус краснухи, вирусы гепатита B и C, цитомегаловирус, энтеровирусы).
Сохранение вируса как биологического вида обеспечивает его восприимчивый хозяин, который является основным элементом экологической ниши вируса. Способность клеток или организма хозяина заражаться называется восприимчивостью.
5-я стадия называется эклипс-фазой, которая характеризуется исчезновением вириона, поскольку он перестает обнаруживаться при электронной микроскопии. В эту стадию начинается синтез компонентов вириона, т.е. его репродукция. Она носит раздельный характер, поскольку компоненты вириона синтезируются в разных частях клетки: белки на рибосомах, нуклеиновые кислоты в ядре или цитоплазме. Вирус использует для этого генетический аппарат клетки, подавляя необходимые ей самой синтетические реакции.
Эта стадия начинается с транскрипции и репликации вирусного генома. Транскрипция вирусного генома двунитевых ДНК-содержащих вирусов происходит, так же как и клеточного генома, по триаде ДНК- иРНК- белок. Различия касаются только происхождения фермента ДНК-зависимой РНК-полимеразы, необходимой для данного процесса. У вирусов, геном которых транскрибируется в цитоплазме клетки хозяина (например, вирус оспы), имеется собственная вирусспецифическая РНК-полимераза. Вирусы, геномы которых транскрибируются в ядре (папова- и аденовирусы, вирусы герпеса), используют содержащуюся там клеточную РНК-полимеразу II или III.
У РНК-содержащих вирусов транскрипция их генома осуществляется несколькими путями:
1. Вирусы с негативным геномом (минус-нитевые), к которым относятся орто-, парамиксо- и рабдовирусы, имеют в своем составе вирусспецифическую РНК-полимеразу или транскриптазу. Они синтезируют иРНК на матрице геномной РНК. Подобный фермент отсутствует в нормальных клетках, но синтезируется клетками, зараженными вирусами. Он находится в составе как однонитевых, так и двунитевых РНК-содержащих вирусов.
2. У вирусов с положительным геномом (плюс-нитевые), к которым относятся пикорна-, тогавирусы и др., функцию иРНК выполняет сам геном, который транслирует содержащуюся в нем информацию на рибосомы клетки хозяина.
3. Особняком стоит группа РНК-содержащих ретровирусов, в составе которых имеется обратная транскриптаза, или ревертаза. Уникальность этого фермента состоит в его способности переписывать информацию с РНК на ДНК. Этот процесс называется обратной транскрипции.
Как отмечалось выше, количество генов в вирусном геноме весьма ограничено. Поэтому для увеличения количества вирусной информации существует своеобразный трансляционный механизм, функционирующий через иРНК, который передает значительно больше информации, чем записано в вирусной нуклеиновой кислоте. Это достигается разными путями, например при транскрипции информации с переписывающихся участков ДНК на иРНК путем сплайсинга (вырезание бессмысленных кодонов и сшивание концов), а также при считывании антикодонами тРНК одной и той же молекулы иРНК с разных нуклеотидов. При этом образуются новые триплеты, увеличивающие количество транслируемой информации.
Вирус-специфическая информация транслируется на рибосомы клетки хозяина, которые предварительно освобождаются от клеточных белков и собираются в вирус-специфические полисомы.
6-я стадия - сборка вириона - состоит прежде всего в образовании нуклеокапсидов. Поскольку синтез вирусных нуклеиновых кислот и белков в клетке происходит в разных ее структурах, необходима транспортировка составных частей вириона в одно место сборки. При этом вирусные белки и нуклеиновые кислоты обладают способностью узнавать и самопроизвольно соединяться друг с другом. В основе самосборки простых вирионов лежит способность вирусных полипептидов соединяться в капсомеры, которые, располагаясь вокруг осей симметрии, образуют многогранник. В других случаях полипептиды в виде спирали окружают вирусную нуклеиновую кислоту.
Многие простые вирионы собираются на репликативных комплексах мембранах эндоплазматического ретикулума. У сложных вирионов сборка нуклеокапсида начинается на репликативных комплексах, а затем продолжается на плазматической мембране, с наружной стороны которой располагаются суперкапсидные гликопротеиды. Затем гликопротеидные и примыкающие к ним с другой стороны нуклеокапсидные участки выпячиваются через клеточную мембрану, образуя почку, как это имеет место у орто- и парамиксовирусов, рабдовирусов. После отделения почки, содержащей нуклеокапсид и суперкапсидные белки, образуются свободные вирионы. Они либо через клеточную плазматическую мембрану проходят во внеклеточное пространство, либо через мембрану эндоплазматического ретикулума проникают в вакуоль эндоплазматической сети. При этом мембранные липиды обволакивают почку, вытесняя из нее белки. Многие ДНК-содержащие вирусы, например вирус герпеса, собираются в ядре клетки на ее мембране, где образуются нуклеокапсиды. Затем они отпочковываются в перинуклеарное пространство, приобретая внешнюю оболочку. Дальнейшее формирование вириона происходит в мембранах цитоплазматического ретикулума и в аппарате Гольджи, откуда вирус транспортируется на поверхность клетки.
Существуют следующие общие принципы сборки вирусов, имеющих разную структуру:
1. формирование вирусов является многоступенчатым процессом с образованием промежуточных форм;
2. сборка просто устроенных вирусов заключается во взаимодействии молекул вирусных нуклеиновых кислот с капсидными белками и образовании нуклеокапсидов (например, вирусы полиомиелита). У сложно устроенных вирусов сначала формируются нуклеокапсиды, с которыми взаимодействуют белки суперкапсидных оболочек (например, вирусы гриппа);
3. формирование вирусов происходит не во внутриклеточной жидкости, а на ядерных или цитоплазматических мембранах клетки;
4. сложно организованные вирусы в процессе формирования включают в свой состав компоненты клетки-хозяина (липиды, углеводы).
7-я стадия - выход вирусных частиц из клетки - происходит двумя путями.
Первый тип — взрывной — характеризуется одновременным выходом большого количества вирусов. При этом клетка быстро погибает. Такой способ выхода характерен для вирусов, не имеющих суперкапсидной оболочки.
Время, необходимое для осуществления полного цикла репродукции вирусов, варьирует от 5—6 часов (вирусы гриппа, натуральной оспы и др.) до нескольких суток (вирусы кори, аденовирусы и др.). Образовавшиеся вирусы способны инфицировать новые клетки и проходить в них указанный выше цикл репродукции.
Простые вирусы, лишенные суперкапсида, например пикорнавирусы, аденовирусы и др., вызывают деструкцию клетки и попадают во внеклеточное пространство. Другие вирусы, имеющие липопротеидную внешнюю оболочку, выходят из клетки путем почкования, в результате чего в течение длительного времени она сохраняет свою жизнеспособность. Такой путь характерен для вируса гриппа и др.
Интегративная инфекция. Интеграция (встраивание) вирусной нуклеиновой кислоты в клеточный геном.
Данный путь взаимодействия между вирусом и клеткой хороша не одинаков для ДНК- и РНК-содержащих вирусов. В первом случае вирусная ДНК в кольцевой форме интегрирует в клеточный геном. При этом место интеграции определяется гомологичными нуклеотидными последовательностями, имеющимися в определенных участках - ДНК сайтах при участии ряда ферментов: рестриктаз, эндонуклеаз, лигаз. Вирус, интегрированный в клеточный геном, называют провирусом.
Провирус может реплицироваться в составе клеточного генома пропорционально делению клетки. При этом каждая дочерняя клетка получает копию провирусного генома. В другом случае амплификация провирусной ДНК с увеличением числа копий провируса без его выщепления из клеточного генома может привести к встраиванию провируса в другую хромосому. Выщепление провируса из клеточного генома и его проникновение в новую клетку может вызвать продуктивную инфекцию.
В случае РНК-содержащих вирусов включение РНК в клеточный геном происходит путем обратной транскрипции. Механизм обратной транскрипции состоит в первоначальном образовании ДНК-транскрипта на матрице РНК при обязательном участии обратной транскриптазы. Этот транскрипт представляет собой одну нить ДНК, являющуюся матрицей для образования второй нити. Затем образовавшийся двунитевои ДНК-транскрипт замыкается в кольцо и встраивается в клеточный геном. Данный процесс объединения вирусной нуклеиновой кислоты с хромосомой клетки хозяина называется вирогениеи. В интегрированном состоянии вирусная ДНК может транскрибироваться в составе клеточного генома при участии клеточных РНК-полимераз.
Биологический смысл интегративного типа взаимодействия между вирусом и клеткой хозяина можно видеть прежде всего в сохранении вирусной информации в составе клеточного генома и ее передаче потомству. Вместе с тем это в определенной степени отражается и на эволюции некоторых вирусов (например, бактериофагов), которые при выщеплении из состава клеточной хромосомы могут захватывать отдельные ее гены.
С другой стороны, подобный тип взаимодействия может отразиться на судьбе клеток хозяина в зависимости от расположения локуса, в котором происходит интеграция вирусного генома, вплоть до расстройства регуляции синтеза белка и неконтролируемого деления клетки. Это может привести к онкогенной трансформации клеток хозяина и развитию разнообразных опухолей.
Дефектные вирусы.
Дефектные вирусы - вирусы, утратившие в процессе репродукции часть своего генома:
- дефектные интерферирующие частицы представляют собой вирионы, содержащие только часть генетической информации исходного вируса; репродуцируются только при участии родственного им вируса-помощника;
- вирусы-спутники отличаются для своей репродукции требуют участия любого вируса-помощника, не обязательно родственного исходному вирусу. например, вирус гепатита D (дельта) репродуцируется только в присутствии вируса гепатита В;
- интегрированные геномы представляют собой провирусы, т. е. вирусные геномы, встроенные (интегрированные) в хромосому клетки хозяина при интегрированной инфекции, которые потеряли способность превращаться в полноценный вирус;
- псевдовирионы - вирионы, имеющие нормальный капсид, содержащий часть собственной нуклеиновой кислоты и фрагменты нуклеиновой кислоты своего хозяина, либо часть хромосомы клетки хозяина и часть нуклеиновой кислоты другого вируса.
Значение дефектных вирусов состоит в их способности переносить генетический материал из клетки донора в клетку реципиента.
Читайте также:
