
Ортомиксовирусы что это такое
Тема занятия: Ортомиксовирусы. Вирус гриппа. Биологические свойства. Патогенез и клиника гриппа. Лабораторная диагностика. Специфическая профилактика гриппа.
Учебная цель занятия: Изучение общих характеристик ортомиксовирусов, биологических свойств вируса гриппа, патогенеза и клиники гриппа, методов лабораторной диагностики и специфической профилактики гриппа.
Задачи занятия: Познакомиться с основными свойствами ортомиксовирусов, биологическими свойствами вируса гриппа, патогенезом и клиникой вызываемого им заболевания, методами лабораторной диагностики и средствами специфической профилактики гриппа.
Общая характеристика ортомиксовирусов
Семейство Orthomyxoviridae (от греч. orthos - правильный, прямой и myxa - слизь) объединяет группу вирусов, которые имеют особое сродство к мукополисахаридам и гликопротеидам клеточных рецепторов, а также сходные биологические свойства (способность агглютинировать эритроциты, тропизм к органам дыхания, легкость культивирования в куриных эмбрионах, наличие гемагглютинина, а у некоторых вирусов – также и нейраминидазы).
Классификация. Согласно международной таксономии вирусов, семейство Orthomyxoviridae включает четыре рода: Influenzavirus А, Influenzavirus В, Influenzavirus С и Togotovirus.
Род Influenzavirus А включает один вид - вирус гриппа А. Этот вирус открыли В. Смит, С. Эндрюс и П. Лейдлоу в 1933 г. Внутри этого вида различают субтипы вируса по гемагглютинину и нейраминидазе. Внутри субтипов могут быть сероварианты. Номенклатура вновь выделенных изолятов вируса гриппа А складывается из следующих критериев: род / хозяин / место выделения / обозначение штамма / год выделения / формула (тип гемагглютинина Н и нейраминидазы N), например, А/крачка/ Южная Африка/1/61/(Н5N3).
Род Influenzavirus В включает один вид - вирус гриппа В, который выделили Т. Фрэнсис и Р. Меджилл в 1940 г. У него в отличие от вируса гриппа А не установлены антигенные субтипы. Вирусы гриппа В обозначаются по следующим критериям: род / место выделения / обозначение штамма / год выделения.
Род Influenzavirus С также включает один вид - вирус гриппа С, который открыл Р. Тэйлор в 1947 г. Данный вид инфицирует только людей. Антигенных субтипов у вируса гриппа С не отмечено. Чаще всего вирусы гриппа С вызывают инфекцию у детей в виде отдельных спорадических вспышек.
Род Togotovirus включает 2 вида, из которых вирус Togoto является прототипным. Тоготовирусы передаются между позвоночными через клещей.
Морфология вирионов. Геном ортомиксовирусов представлен односпиральной линейной фрагментированной молекулой минус-РНК. Количество фрагментов в геноме различных представителей ортомиксовирусов разное. Так, вирусы гриппа А и В имеют 8 фрагментов, вирусы гриппа С – 7 фрагментов; тоготовирусы - 6 фрагментов. Каждый фрагмент в отдельности и все фрагменты в совокупности покрыты белковой оболочкой, образуя нуклеокапсид. Снаружи вирион ортомиксовирусов покрыт суперкапсидной оболочкой, имеющей шипы, образованные гемагглютинином и нейраминидазой. Между нуклеокапсидом и суперкапсидом располагается матриксный белок М. Нуклеокапсид с М-белком в совокупности составляют сердцевину вириона.
Устойчивость. Ортомиксовирусы слабо устойчивы во внешней среде. Под действием ультрафиолетового света они погибают в течение 5 минут. В 70%-ном этиловом спирте вирусы гибнут через 5 минут, в 3%-ном феноле, 1%-ной настойке йода, 1%-ной сулеме, 1%-ном купоросе – через 3 минуты.
Морфология. Вирионы вируса гриппа являются “одетыми”, сферической или нитевидной формы, полиморфные, размером 80-120 нм, содержат 32 капсомера. Снаружи они имеют липопротеиновую оболочку, которая образует выступы (шипы). Вирионы ортомиксовирусов состоят из нуклеокапсида спиральной симметрии и липопротеидной оболочки (суперкапсида), которая имеет выступы, образованные из гемагглютинина и нейраминидазы. Между нуклеокапсидом и суперкапсидом располагается М-белок.
Геном вирионов вируса гриппа образован однонитевой спиральной молекулой минус-РНК. Геном состоит из отдельных фрагментов (у вирусов гриппа А и В – 8 фрагментов, у вируса гриппа С – 7 фрагментов). Каждый фрагмент вирусной РНК покрыт отдельным белком. РНК упакована в комплексе с белками в виде спирали, составляя кор или нуклеокапсид. На поверхности вирионов имеется множество мелких шипов, построенных из гликопротеинов с гемагглютинирующей или нейраминидазной активностью.
Нуклеокапсид имеет спиральный тип симметрии. Он представляет собой рибонуклеопротеиновый (РНП) тяж (РНК, связанная с капсидными белками), уложенный в виде двойной спирали, которая составляет сердцевину вириона. Сердцевина окружена мембраной, состоящей из белка М, который соединяет РНП с двойным липидным слоем внешней оболочки (суперкапсидом) и шиповидными отростками гемагглютинина и нейраминидазы.
Суперкапсид образован липидным бислоем, который пронизывают гликопротеиновые шипы (спикулы), определяющие гемагглютинирующую (Н) или нейраминидазную (N) активность. В вирионе обнаружено 7 структурных белков, из которых 3 белка являются наружными и 4 белка - внутренние.
К наружным структурным протеинам относятся гемагглютинин (Н), нейраминидаза (N) и белок М. Гемагглютинин и нейраминидаза входят в состав суперкапсида. Они заякорены гидрофобными концами в липидном бислое суперкапсида и структурно оформлены в виде шипов, выступающих на поверхности вириона. Внутри вириона гемагглютинин и нейраминидаза контактируют с М-белком, а через него - с сердцевиной.
Гемагглютинин и нейраминидаза являются гликопротеинами и входят в состав липопротеиновой оболочки вириона (суперкапсида). Гемагглютинин изменчив (у вирусов гриппа А человека и животных выявлено 16 антигенных типов или вариантов гемагглютинина – от Н1 до Н16). Антитела к нему обеспечивают защитный эффект. Функции гемагглютинина: - прикрепительная (связывается с сиаловой кислотой рецепторов поверхности эпителиальных клеток); - участие в слиянии (содержит пептид слияния) оболочки вириона с мембранами клетки и гемагглютинации, агрегируя (склеивая) эритроциты; - протективные свойства, способствующие развитию иммунитета (вируснейтрализующие антитела).
Нейраминидаза распознает и взаимодействует с рецепторами клетки, содержащими N-ацетилнейраминовую кислоту, то есть приводит к проникновению вируса внутрь клетки, а также, отщепляя нейраминовую кислоту от дочерних вирионов и клеточной мембраны, к выходу вирусов из клеток и распространению вирионов. Нейраминидаза изменчива – у вируса гриппа А человека и животных выявлено 9 вариантов нейраминидазы (от N1 до N9). У вируса гриппа типа С нейраминидаза отсутствует; ее заменяет ацетилэстераза, отрезающая ацетильные группы гликопротеинов. Нейраминидаза образует отдельные шипы, которых меньше, чем количество шипов гемагглютинина. Нейраминидаза отделяет вирионы от сиалированных муцинов, покрывающих слизистую оболочку, способствуя продвижению вируса к поверхности эпителиальных клеток. При завершении репликативного цикла она помогает отделению созревших вирионов от эпителиоцитов.
Протеин М (матриксный белок) выстилает липопротеиновую оболочку изнутри вирусной частицы. Он представлен белками М1 и М2. Матриксный белок М1 и мембранный белок М2 окружают нуклеокапсид и защищают геном. При этом М1-белок взаимодействует с нуклеокапсидом и оболочкой, а М2-белок формирует мембранный канал.
Три внутренних структурных протеина (РВ1, РВ2, РА) формируют нуклеокапсид, а также участвуют в транскрипции и репликации вируса. Так, белки Р1 или РВ1 являются транскриптазой, Р2 или РВ2 – эндонуклеазой, Р3 или РА – репликазой. Четвертый внутренний белок (NР) связан с наружным фрагментом РНК и выполняет регуляторные и структурные функции. Кроме структурных белков для вируса гриппа характерны неструктурные белки NS1и NS2, существующие только в инфицированной клетке.
Репродукция вируса гриппа состоит из следующих этапов: 1. Адсорбция и проникновение вируса внутрь клетки. Вирус адсорбируется на клетках за счет взаимодействия гемагглютинина с сиаловой кислотой рецепторов клеточных мембран и проникает внутрь.
2. Депротеинизация капсида и его проникновение в ядро клетки. В цитоплазме клетки нуклеокапсид освобождается от суперкапсида и выходит в цитоплазму. Отсюда вместе с М-белком он транспортируется в ядро, где уже через несколько минут после заражения появляются первые РНК-транскрипты.
3. Образование полных и неполных плюснитей РНК. В ядре клетки происходит трансформация геномной минус-нити РНК с помощью вирусной РНК-зависимой РНК-полимеразы в неполные и полные плюс-нити. Неполные плюс-нити являются иРНК для синтеза вирусных белков, а полные плюс-нити служат матрицей для синтеза геномных минус-нитей РНК.
4. Трансляция вирусных белков. После выхода молекул мРНК из ядра в цитоплазму клетки происходит трансляция вирусных белков (NP, РВ1, РВ2, РА и М) на полирибосомах. Белки NP, РВ1, РВ2 и РА после синтеза из цитоплазмы возвращаются в ядро, где связываются с вновь синтезированной вирусной РНК, а затем в виде нуклеокапсида возвращаются в цитозоль. Матриксный белок М после синтеза в цитоплазме движется к внутренней поверхности клеточной мембраны, вытесняя из нее в этом участке клеточные белки. Белки N и H синтезируются на рибосомах, связанных с мембранами эндоплазматического ретикулума, подвергаются гликозилированию и транспортируются на внешнюю поверхность клеточной мембраны, встраиваясь напротив белка М и образуя шипы. Белок Н при этом подвергается разрезанию на субъединицы НА1 и НА2.
5. Формирование потомства вирионов. На границе ядерной мембраны белки капсида и РНК потомства формируют нуклеокапсиды, которые мигрируют в цитоплазму для дальнейшего созревания. В модифицированных М-белком участках клеточной мембраны нуклеокапсиды окончательно созревают и выходят из клеток путем почкования.
6. Выход вирионов из клетки происходит почкованием. При этом сформированный нуклеокапсид, проходя через мембрану клетки, окружается белком М и измененной мембраной клетки, содержащей гемагглютинин и нейраминидазу.
Историческая справка. Слово “грипп” заимствовано из французского языка и в понятии “эпидемический (заразный) насморк” используется в России с начала XIX в. Значительно раньше болезнь называли “инфлюэнцей”. Термин “инфлюэнца” принят во многих странах, но у нас утвердилось понятие “грипп”, отражающее клинико-эпидемиологическую сущность болезни: острое начало, быстрое развитие и распространение среди людей (фр. grippe - хватать, схватывать). Первая из наиболее полно описанных эпидемий гриппа относится к 1510 г. В 1918 г. случилась необычная по масштабам и последствиям пандемия (глобальная эпидемия) гриппа. Как и большинство крупных вспышек, она возникла в Юго-Восточной Азии (Китае), но по случайному стечению обстоятельств зафиксирована в истории как “испанский грипп”, или просто “испанка”. В течение 2-3 лет от испанки погибло более 20 млн. человек. Последующие эпидемии гриппа, хотя и перерастали неоднократно в пандемии, не приносили столько жертв.
Современная история гриппа началась в 1933 г., когда был выделен его возбудитель – вирус гриппа типа А. Это удалось сделать В. Смиту и его коллегам при заражении африканских хорьков ультрафильтратом смыва из зева больного гриппом (им был В. Смит - один из авторов открытия). Заболевание, которое последовало у зараженных животных, очень напоминало грипп человека. В 1940 г. был изолирован вирус гриппа типа В, а в 1947 г. – вирус гриппа типа С.
Экология. Кроме человека в естественных условиях вирусы гриппа инфицируют млекопитающих и птиц. Вирусы гриппа человека не вызывают эпизоотии, хотя пример с “испанским гриппом” свиней не исключает такой возможности. Так, в последние годы зарегистрированы случаи заражения людей штаммами вируса гриппа А, циркулирующими среди птиц. Реальная тревога возникла в 1997 г. в Гонконге, когда 18 человек напрямую заразились от домашних птиц вирусом Н5N1; 6 человек погибло. В 2003 г. высоковирулентные штаммы птиц Н5N1 и Н7N7 спровоцировали фатальные случаи гриппа среди жителей Южной Азии. Вирусы типов В и С являются сугубо человеческими и в естественных условиях не инфицируют других хозяев.
Патогенез. Вирусы гриппа проникают в респираторный тракт с каплями аэрозоля и частицами пыли. Чем мельче их величина, тем глубже проникает вирус; самые мелкие частицы достигают бронхиол и альвеол. Главным местом размножения вируса служат реснитчатые и бокаловидные клетки слизистой оболочки верхних дыхательных путей, но инфекция может охватить весь респираторный тракт - от носовых ходов до терминальных бронхиол и альвеол. Повреждение зараженных эпителиоцитов приводит к выходу вируса в кровь. Находясь в крови, вирус гриппа не оказывает патологического воздействия из-за отсутствия сывороточных и тканевых протеаз, способных активировать вирусный гемагглютинин.
В зонах выраженного повреждения эпителия могут развиваться осложнения, вызванные присоединением бактериальной инфекции.
Освобождение от вируса начинается после повреждения слизистой оболочки. Продукты распада эпителиоцитов и медиаторы воспаления стимулируют продукцию слизи, а уцелевшие клетки мерцательного эпителия и кашлевые толчки проталкивают ее вверх вместе с основной массой вируса. Этот простой, но надежный механизм в сочетании с ринореей предупреждает распространение инфекции по респираторному тракту, способствуя выздоровлению.
Вирус гриппа А. Эпидемиология. Источник инфекции - только человек, больной или носитель. Заражение происходит воздушно-капельным путем. Инкубационный период очень короток, возникшая эпидемия распространяется очень быстро и при отсутствии коллективного иммунитета может перерасти в пандемию.
Особенности патогенеза и клиники. Инкубационный период при гриппе короткий - 1-2 суток. Вирус размножается в эпителиальных клетках слизистой оболочки дыхательных путей с преимущественной локализацией в области трахеи, что клинически проявляется в виде сухого мучительного кашля с болями по ходу трахеи. Продукты распада пораженных клеток попадают в кровь, вызывают сильную интоксикацию и повышение температуры тела до 38-39°С. Вирус гриппа оказывает угнетающее действие на кроветворение и иммунную систему. Все это может приводить к вторичным вирусным и бактериальным инфекциям, которые осложняют течение болезни.
Постинфекционный иммунитет. Главная роль в формировании приобретенного иммунитета принадлежит вируснейтрализующим антителам, блокирующим гемагглютинин и нейраминидазу, а также секреторным иммуноглобулинам IgAs. Перенесенное заболевание оставляет за собой антитела, которые сохраняются долгие годы. Связанный с этими антителами иммунитет имеет штаммовую направленность и не гарантирует защиты от заражения новыми вариантами вируса.
Лабораторная диагностика. Материалом для исследования служит отделяемое носоглотки, которое получают либо путем смыва, либо с помощью ватно-марлевых тампонов, и кровь. Для диагностики гриппа применяют следующие методы: 1. Вирусологический - заражение куриных эмбрионов, культур клеток почек зеленых мартышек (Vero) и собак (МDСК). Культуры клеток особенно эффективны для выделения вирусов А (Н3N2) и В. 2. Серологический - выявление специфических антител и определение возрастания их титра (в парных сыворотках) с помощью РТГА, РСК, иммуноферментного метода. 3. В качестве ускоренной диагностики используют иммунофлуоресцентный метод, позволяющий быстро обнаружить вирусный антиген в мазках-отпечатках со слизистой оболочки носа или в смывах из носоглотки больных. 4. Разработаны методы иммуноферментного анализа для выявления вирусных антигенов непосредственно в материале от больных, а также метод РНК-зонда. 5. Предложена иммуноферментная методика “бескровного” обнаружения специфических секреторных IgAs в слюне в качестве самостоятельного способа диагностики гриппа.
Вирус гриппа В. Структура вириона сходна со структурой вируса гриппа типа А. Геном состоит из 8 фрагментов, кодирующих 3 неструктурных и 7 структурных белков. По антигенным свойствам гемагглютинина и нейраминидазы различают несколько серовариантов. Процесс антигенного дрейфа выражен слабее, чем у вируса типа А. Вирусы гриппа типа В вызывают локальные вспышки и эпидемии; пандемий не вызывают. Клиника заболевания такая же, как и при гриппе типа А. Лабораторная диагностика такая же, вирус дифференцируется серологически. Специфическая профилактика осуществляется так же, как против гриппа А.
Вирус гриппа С. Вирион типа С имеет такую же форму, как вирусы типов А и В. Однако он отличается от них не только антигенными свойствами, но и рядом других признаков. Геном представлен однонитевой негативной РНК из 7 фрагментов, нуклеотидная последовательность которых существенно отличается от таковых вирусов типов А и В. Геном кодирует 1-2 неструктурных и 6 структурных белков. У вируса типа С отсутствует нейраминидаза, поэтому на наружной мембране вириона имеется только один тип шипов.
Вирусу типа С не свойственна такая изменчивость, как вирусу типа А. Хотя вирус гриппа С не вызывает пандемий и больших эпидемий, он часто является причиной спорадических заболеваний гриппом. Клиника заболевания такая же, как при относительно умеренных формах гриппа А. Диагностика основана на выделении вируса в куриных эмбрионах; применяются также иммунофлуоресцентный метод и другие серологические реакции.
Лечение гриппа у людей проводится с использованием амантадина, ремантадина, противогриппозного иммуноглобулина, интерферона.
Специфическая профилактика заключается в применении вакцин: - живая из аттенуированного вируса; - убитая вакцина цельновирионная; - субвирионная вакцина (из расщепленных вирионов); - субъединичная вакцина (вакцина, содержащая только гемагглютинин и нейраминидазу). Полимер-субъединичная вакцина “Гриппол” представляет собой конъюгат поверхностных белков вирусов гриппа А и В, который связан с сополимером полиоксидионием (иммуностимулятор).
Контрольные вопросы по теме занятия: 1. Общая характеристика ортомиксовирусов. 2. Строение вириона вируса гриппа. 3. Репродукция вируса гриппа. 4. Эпидемиология и клиника гриппа. 5. Лабораторная диагностика гриппа. 6. Специфическая терапия и профилактика гриппа.
Литература для подготовки к занятию: Основная литература: 1. Медицинская микробиология, вирусология и иммунология. Под ред. А.А. Воробьева. М., 2004. Дополнительная литература: 1. Л.Б. Борисов. Медицинская микробиология, вирусология, иммунология. М., 2002. 2. О.К. Поздеев. Медицинская микробиология. М., ГЭОТАР-МЕДИА, 2005. 3. Медицинская микробиология. Справочник. Под ред. В.И. Покровского и О.К. Поздеева. М., ГЭОТАР-МЕД, 1998.
Таблица 2. Различие в свойствах
ортомиксовирусов и парамиксовирусов
Сферическая, нити в свежем изоляте (выделенном вирусе)
Сегментированный, восемь частей РНК
Цельная линейная молекула РНК
Место синтеза рибонуклеопротеида
ДНК-зависимый синтез РНК
Необходим для размножения
Эффект актиномицина Д
вызывая гемагглютинцию. Название напоминало сродство вирусов к муцинам (от греч. myxa, означающее слизь). В эту группу были включены вирусы гриппа, свинки (паротита), Ньюкаслской болезни и парагриппа. Последующее обнаружение важных различий между вирусами гриппа и другими вирусами в этой группе привело к их переклассификации на два отдельных семейства: Orthomyxovirdae, состоящего из вирусов гриппа и Paramyxoviridae, состоящего из вируса болезни Ньюкасл, вируса паротита, вирусов парагриппа, кори и респираторно-синцитиального вируса. Таблица 1 перечисляет важнейшие различия между ортомиксовирусами и парамиксовирусами.
2. Ортомиксовирусы Ортомиксовирусы
Это - большая группа вирусов (приблизительно 20 типов) обладающих сродством к мукопротеинам (отсюда и название, греч.. myxa слизь).
Они вызывают грипп у человека, грипп животных (свиней, лошадей), и грипп и чуму птиц.
Ортомиксовирусы имеют сферическую форму и размеры 80-120 нм. Нуклеокапсид характеризуется спиральной симметрией. Вирионы состоят из нуклеоида в форме спирали нуклеопротеида и внешней липо-углеводо-протеидной мембраны и содержат фермент, который разрушает мукопротеиновые рецепторы клеток восприимчивых людей и животных. Вирионы чувствительны к эфиру и дезксихолату натрия.
Вирусная этиология гриппа типа А была установлена в 1933 г. У. Смитом, К. Эндрюсом и П. Лэйдлоу. Они установили, что профильтрованные смывы из носоглотки больных гриппом на ранних стадиях болезни были инфекционными. В 1940 г. Т. Френсис и Т. Меджилл открыли вирус гриппа В, а в 1947 г. Р. Тейлор - вирус гриппа С.
Морфология и химический состав. Вирусы гриппа имеют округлую или овальную форму, диаметр 80 -100 нм.
Нуклеокапсид образован рибонуклеопротеидом, покрытым сверху наружной липидно-углеводно-протеиновой оболочкой (суперкапсид).
Белки вируса состоят из семи отличающихся друг от друга полипептидов, четыре из которых связаны с нуклеокапсидом, а три - с наружной оболочкой. Липиды и углеводы взаимосвязаны с вирусными белками, их синтез и специфичность зависят от генома клетки.
Геном вируса представлен однонитчатой РНК, фрагментированной у вирусов гриппа А и В на восемь, вируса гриппа С – на семь фрагментов. Фрагменты в составе вириона слабо связаны между собой.
В составе вириона также присутствует вирусная РНК-зависимая РНК-полимераза: она необходима для инвазионной способности, так как вирионная РНК является минус-нитевой и поэтому должна быть транскрибирована для образования вирусной мессенджер – РНК (мРНК). Вирион окружен оболочкой, содержащей липиды, заимствованные из клеточной мембраны. От вторичной оболочки выступают шипы (рис. 1), которые обеспечивают прикрепление вириона к рецепторам клетки, в результате они способны склеивать эритроциты некоторых видов животных и называются поэтому гемагглютинином (H). Они имеют длину примерно 10 нм и состоят из тримеров идентичных гликопротеидных субъединиц, каждая из которых состоит из двух полипептидных цепей, HA1 и HA2. Эти два полипептида связаны в каждой субъединице одной дисульфидной связью. В вирусной частице молекула гемагглютинина связана с липидной мембраной гидрофобным участком вблизи карбоксильного конца HA2. Вирусы гриппа прикрепляются к клеткам с помощью гемагглютинина, взаимодействующего с мембранными рецепторами, содержащими N-ацетилнейраминовую кислоту (NANA).
Между гемагглютиновыми шипами имеются грибовидные выступы нейраминидазы (N). Головка имеют коробчатую форму и собрана из четырех приблизительно шаровидных субъединиц, присоединенных к стеблю, содержащему гидрофобный регион, встроенный в вирусную оболочку. Фермент катализирует расщепление NANA и прилежащего углеводного остатка в гликопротеине слизи.
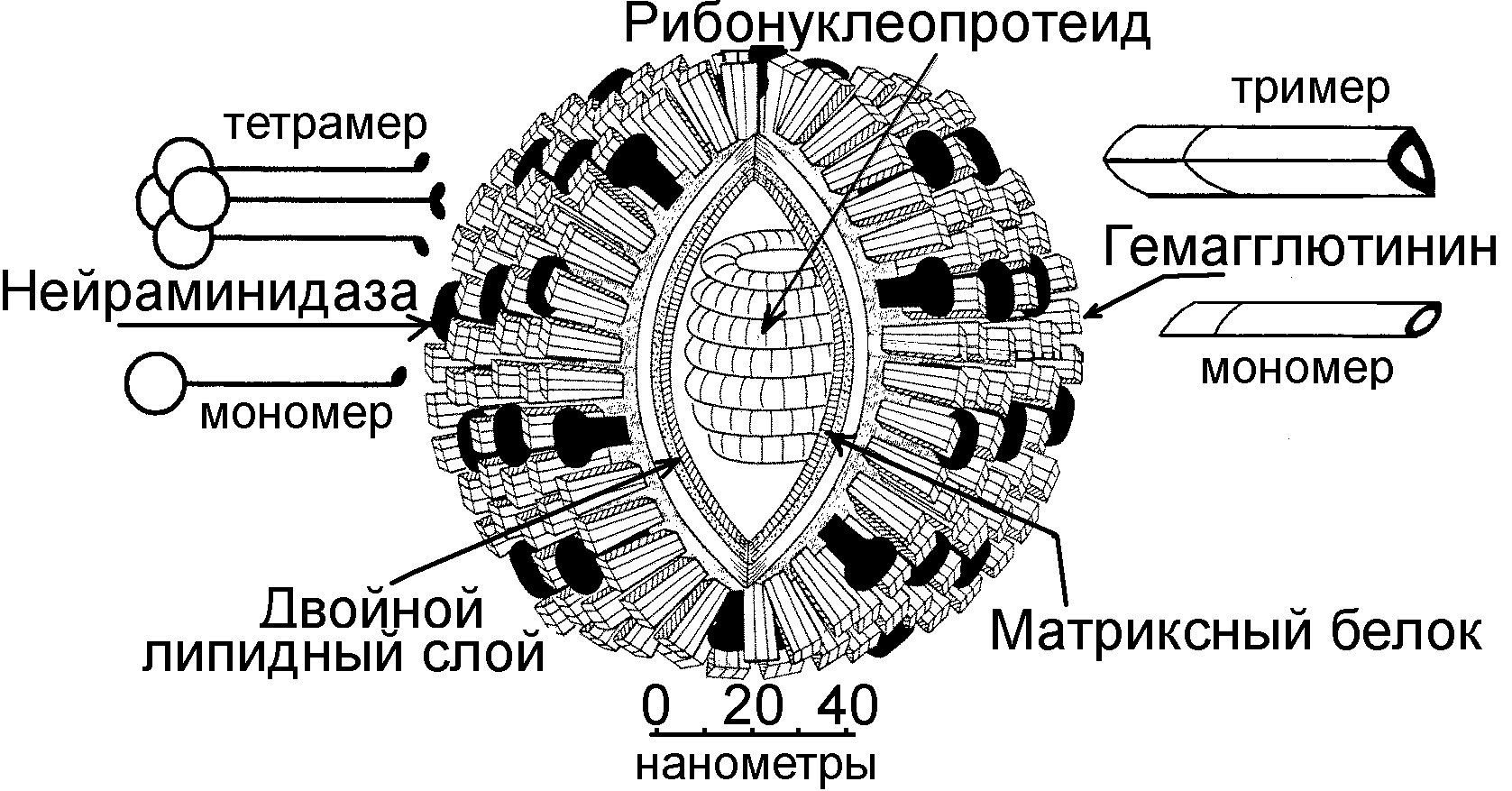
Рис. 1. Схематическое изображение вириона гриппа
NANA всегда присутствует в недавно синтезированных вирионах и ее удаление нейраминидазой предотвращает агглютинацию новых вирусных частиц, что ведет к увеличению числа свободных вирусных частиц и, следовательно, распространению вируса из первоначального очага инфекции.
Вирусы гриппа А и В имеют гемагглютинин и нейраминидазу, вирус гриппа типа С – только гемагглютинин.
ОРТОМИКСОВИРУСЫ — семейство Orthomyxoviridae РНК-содержащих пневмотронных вирусов, включающее вирусы гриппа А, В и С. В род вирусов гриппа А входят вирусы гриппа А человека, свиней, лошадей и птиц. Вирусы гриппа В и С поражают только человека.
В 1955 г. К. Эндрюс и др. объединили в одну группу вирусы, имеющие РНК-геном, спиральную симметрию нуклеокапсида, полиморфную липопротеидную оболочку, обладающие способностью к гемагглютинации и ферментной элюции вследствие расщепления нейраминидазой мукопротеидов оболочки эритроцитов. Они дали нм название миксовирусов. В 1962 г. Уотерсон (А. P. Waterson) выделил в группе миксовирусов две подгруппы, получившие в дальнейшем статус семейств,— ортомиксовирусы ипарамиксовирусы (см.).
Форма вирионов О. сферическая (рис. 1, а) или несколько вытянутая, диам. 80—120 нм; встречаются нитевидные формы (рис. 1,6) шириной 80—100 нм и длиной до 4000 нм, особенно у свежевыделенных штаммов. Мол. масса сферических вирионов 2,5 X 10 8 дальтон, коэффициент седиментации 722—800S, плавучая плотность вирусов гриппа А и В — 1.19 г/см 3 . гриппа С — 1,17— 1,19 г/см 3 . Белок составляет 60—75% веса вириона, липиды — 18—37%, углеводы — 5—9%, РНК — 0,8— 1,1%.
Геном О. представляет собой одноцепочечную линейную РНК, состоящую из 7—8 (по нек-рым данным, из 10) отдельных молекул (фрагментов) с мол. массой до 1 X 10 6 дальтон; общая масса РНК— 4—5 X 10 6 дальтон. Вокруг РНК по спирали располагаются молекулы белка с мол. массой 53 000—65 000 дальтон (примерно 12 молекул на один виток спирали) и небольшое число молекул белка P (вирионной транскриптазы) с мол. массой 81 000—94 000 дальтон.
Отдельные нити нуклеопротеида сложены вдвое в виде шпильки, концы к-рой скручены вместе, вследствие чего образуется двойная спираль с петлей на одном конце (рис. 2). Нуклеопротеид окружен внутренней оболочкой вириона, образованной мембранным белком М с мол. массой 21 000—27 000 дальтон. Снаружи вирион имеет двойной липидный слой с выступами: по составу липидов он идентичен плазматическим мембранам клетки, в к-рой происходила репликация вируса, но не содержит сиаловой к-ты. Ок. 20% массы вириона составляют липиды, из к-рых фосфолипиды составляют 10—13%, холестерин — б—8% , гликолипиды— 1—2%. Клетка-хозяин определяет также углеводный состав вириона: гликолипиды и гликопротеиды содержат галактозу, маннозу, глюкозамин и фукозу.
Расположенные на наружной оболочке вириона выступы имеют длину 10—14 нм и диам. 4—8 нм; расстояние между ними 7—8 нм. Выступы образованы гемагглютинином (ГА) и нейраминидазой (НА), относящимися к гликопротеидам. Гемагглютинин имеет мол. массу 75 000—80 000 дальтон и обусловливает адсорбцию вирионов на клеточных рецепторах. Мол. вес нейраминидазы 55 000— 70 000 дальтон. У вируса гриппа С нейраминидаза не обнаружена.
В зараженных О. клетках синтезируются также два неструктурных белка: NSX с мол. массой 20 000— 25 000 дальтон и NS2 с мол. массой 11 000 дальтон, функция их не ясна.
Репликацию О. изучали в основном на модели вируса гриппа А. С помощью гемагглютинина вирус адсорбируется на чувствительных клетках. Рецепторами вируса на поверхности клетки являются гликопротеиды с N-ацетилнейраминовой (сиаловой) к-той. Процесс проникновения вирионов в клетку не ясен: согласно одной точке зрения, они активно целиком поглощаются клеткой, т. е. имеет место виропексис; согласно другой, — липопротеидная оболочка вириона сливается с плазматической мембраной клетки, затем оболочка и мембрана расплавляются, и нуклеопротеид попадает в цитоплазму.
С помощью транскриптазы, входящей в состав вирусного рибонуклеопротеида, синтезируется РНК, комплементарная вирусной, —кРНК; процесс этот продолжается 3—4 часа и происходит в цитоплазме клетки. Каждый фрагмент родительской РНК транскрибируется отдельно, а каждая образующаяся молекула кРНК является моноцист-ронной матрицей, на к-рой синтезируются вирусные бел ки. Синтез вирионной РНК (вРНК) происходит в основном со 2-го по 6—7 час. после инфицирования клеток. Из общего количества синтезируемой РНК 10% составляет кРНК и 90% вРНК.
Вирусные белки синтезируются на полисомах, образованных вирусными к PH К и клеточными рибосомами, а затем мигрируют в различные участки клетки: нуклеопротеид появляется в ядре, белок NS — в ядрышке, гемагглютинин перемещается с шероховатых мембран на гладкие мембраны эндоплазматической сети, где накапливается также белок М. Сразу после синтеза белка гемагглютинина к нему присоединяются углеводы, синтезированные в клетке.
Репликация О. зависит от функционирования клеточного генома, хотя и не ясно, в чем эта связь заключается.
Созревание (сборка) вириона происходит у поверхности клетки. В нек-рых участках плазматической клеточной мембраны ее белки замещаются вирусными полипептидами— гемагглютинином и нейраминидазой. Нейраминидаза удаляет нейрамино-вую к-ту из тех модифицированных участков клеточной мембраны, из к-рых впоследствии формируется внешняя оболочка вирионов; тем самым вирионы лишаются рецепторов, аналогичных рецепторам, имеющимся у клеточной оболочки.
На внутренней поверхности измененной клеточной мембраны формируется белковый слой, состоящий из белка М. К этим участкам присоединяется вирусный рибонуклеопротеид, после чего от клеточной мембраны отпочковываются вирионы.
Пассажи О. при высокой множественности заражения приводят к образованию неполных частиц (феномен Магнуса), содержащих уменьшенное количество или вообще не содержащих крупномолекулярных фрагментов РНК. Эти частицы не инфекционны, но обладают гемагглютинирующей, иммуногенной и интерферирующей активностью (неинфекционный антиген).
Фрагментированность генома вирусов гриппа обусловливает легкость получения рекомбинантов в условиях эксперимента. Генетическая рекомбинация и множественная реактивация (см.Вирусы) возможны только между представителями одного рода. Между членами разных родов О. может происходить только фенотипическое смешение (см.Негенетические взаимодействия вирусов).
Все О., относящиеся к одному роду, имеют общие, специфические для данного рода антигены, к-рыми являются бел ки нуклео протеида и белок М, но различные поверхностные гликопротеиды — гемагглютинин и не йрамини да з а.
Для вирусов гриппа А человека характерно частое изменение и замена гемагглютинина и нейраминидазы. В меньшей степени изменчивость свойственна вирусам гриппа А свиней и лошадей и вирусу гриппа В. Антигенная структура вирусов гриппа А птиц и вируса гриппа С, по-видимому, мало подвержена изменениям.
О. чаще всего выделяют и культивируют в куриных эмбрионах при заражении в аллантоисную и (или) амниотическую полость. Многие штаммы могут размножаться в культурах почечных клеток обезьян, телят, свиней, хомячков, кур, плода человека, однако обычно после предварительной адаптации.
Среди лабораторных животных наиболее чувствительны к вирусам гриппа человека и млекопитающих животных африканские хорьки при заражении через ноc, менее восприимчивы мыши; сирийские хомячки, морские свинки малочувствительны. Вирусы гриппа птиц (высоко патогенные штаммы) вызывают заболевания у кур при разных способах введения, у мышей при инокуляции в мозг или через ноc, а также при интрацеребральном заражении голубей, уток, обезьян резус, молодых кроликов и морских свинок.
Вирусы гриппа А человека являются причиной эпидемий и пандемий гриппа. В естественных условиях эти вирусы могут вызывать заболевание у свиней, собак и нек-рых других животных. Вирусы гриппа В являются причиной эпидемий и эндемий. Вирусы гриппа С обусловливают эндемичные заболевания (обычно у детей) и внутрибольничные вспышки.
Вирусы гриппа А лошадей вызывают у этих животных обычно нетяжелое респираторное заболевание, но нек-рые штаммы отличаются кардиотропизмом. Экспериментально установлена способность вируса гриппа лошадей вызывать легкое респираторное заболевание у человека.
Вирус гриппа свиней вызывает у этих животных острое респираторное заболевание. Оно протекает тяжело, у молодняка нередко с летальным исходом. Случаи заболевания людей гриппом свиней наблюдаются редко и протекают в виде легкой респираторной инфекции. Существует предположение, что в 1918 г. вирус гриппа человека адаптировался к свиньям и превратился в возбудителя гриппа свиней, поскольку ранее у них это заболевание не регистрировалось.
Гриппозная инфекция у птиц протекает крайне разнообразно — бессимптомно, в виде легкого респираторного заболевания, тяжелой диареи или как чума птиц с поражением нервной системы и высокой летальностью. Эпизоотии наблюдаются среди домашних птиц, гл. обр. семейства куриных. Несколько вирусов было выделено от диких птиц, однако заболевания наблюдались редко (у крачек и чаек). Патогенность вирусов гриппа птиц для человека точно не установлена.
Библиография: Вирусы гриппа и грипп, под ред. Э. Д. Кильбурна, пер. с англ., с. 20, и др., М., 1978; Феннер Ф. и др. Биология вирусов животных, пер. с англ., т. 1, с. 154 и др., т. 2, с. 385 и др., М., 1977; D о w d 1 e W. R. a. o. Orthomyxo-viridae, Intervirology, y. 5, p. 245, 1975.
Источник: Большая Медицинская Энциклопедия (БМЭ), под редакцией Петровского Б.В., 3-е издание
Читайте также:
