
У кишечной палочки атф синтезируется
Верны ли следующие суждения
А. Делением размножаются только клетки, но не органоиды.
Б. Клетки размножаются делением. а вирусы нет.
1. Верно только А
2. Верно только Б
3. Верны обе формулировки
4. Обе формулировки не верны
А. При дигибидном скрещивании у гибридов каждая пара признаков наследуется независимо от других и дает с ними разные сочетания.
Б. Это закон чистоты гамет: в каждую гамету попадает только одна аллель из пары аллелей данного гена родительской особи.
Б. Пары альтернативных признаков не смешиваются и при образовании гамет по одному переходят в них в чистом виде.
А. Результатами эволюции можно считать разнообразие организмов и их приспособленность к условиям окружающей среды.
Б. Результатами эволюции считаются изоляция, борьба за существование и возникновение мутаций.
А. Наследственная изменчивость служит материалом для естественного отбора.
Б. Наследственная изменчивость способствует сохранению в популяции наиболее приспособленных особей
А. Многие белки выполняют каталитическую функцию.
Б. Некоторые гормоны имеют белковую природу.
Упрощение в строении животных, связанные с паразитическим образом жизни, относят к биологическому регрессу.
Б. Возникновение класса Насекомые, сопровождающееся повышением нового уровня их организации, - пример ароморфоза.
А. Генетический код триплетен – один триплет всегда кодирует только одну аминокислоту.
Б. Генетический код однозначен – в молекуле нуклеиновой кислоты одна аминокислота кодируется сочетанием трех последовательно расположенных нуклидов.
А. В результате митоза из одной диплоидной клетки образуется 4 гоплоидные.
Б. Редукция числа хромосом происходит в анафазе I .
А. Мутационная изменчивость носит случайный характер.
А. Митоз обеспечивает рост организма, регенерацию, лежит в основе бесполого размножения.
Б. Мейоз обеспечивает образование гамет, увеличивает генетическое разнообразие и лежит в основе комбинативной изменчивости.
Верны ли следующие суждения о фотосинтезе?
А. В световой фазе фотосинтеза происходит синтез Атор, образуются атомы водорода и молекулярный кислород.
Б. Световая фаза фотосинтеза происходит в строме хлоропласта.
Верны ли следующие суждения о химическом составе клетки?
А. Цинк относится к микроэлементам
Б. В состав хлорофилла входит железо.
Верны ли следующие суждения о строении и функциях органоидов?
А. Лизосомы относятся у двумембранным органоидам.
Б. У кишечной палочки АТФ синтезируются не в митохондриях.
Верны ли следующие суждения об обмене веществ?
А. Второй этап катаболизма – неполное окисление или бескислородный – протекает в цитоплазме..
Б. В результате гликолиза из одной молекулы глюкозы образуется 2 молекулы АТФ.
Верны ли следующие суждения об особенностях ферментов?
А. Ферменты – это специфические белки, которые присутствуют во всех живых организмах и играют роль биологических катализаторов.
Б. Ферменты в отличие от химических катализаторов способны катализировать несколько различных реакций.
Верны ли следующие суждения о приспособленности организмов?
А. К этиологическим приспособлениям относятся, н/п иглы ежа, дикобраза, колючки кактуса, акации, барбариса, которые защищают животных от врагов или препятствуют поеданию растений травоядными животными.
Б. Японские макаки зимой при наступлении холодов спускаются с гор к термальным источникам и подолгу греются в теплой воде – это пример физиологической адаптации.
Верны ли следующие суждения о происхождении жизни?
А. Невозможность самозарождения была доказана опытами Ф. Реди, Л. Пастера и др.
Б. Доказательствами инопланетного происхождения жизни служат НЛО, наскальные изображения летательных аппаратов
А. Молекулы ДНК, находящиеся в митохондриях и хлоропластах, не являются хранителями наследственной информации.
Б. Мутации, возникающие в молекулах ДНК, передаются на и-РНК в соответствии с правилами комплементарности.
А. Плазматическая мембрана состоит из лидидного бислоя и встроенных белков.
Б. Мембранные белки выполняют транспортные, рецепторные и другие функции.
А. В разных фазах мейоза клетка может нести диплоидный или гаплоидный набор хромосом.
Б. В течении всего митоза клетка кожи человека всегда диплоидна.
Верны ли следующие суждения:
№ 2: Верны обе формулировки
№ 3: Верно только А (борьба за существование и мутации – это факторы эволюции, а не результат)
№ 4 верно А (Б – неверно, т.к. сохранению в популяции наиболее приспособленных особей способствует естественный отбор, а наследственная изменчивость представляет собой материал для отбора).
№ 5 верны оба суждения
№ 6 Верно Б (А – неверно, т.к. упрощение в строении животных, связанное с паразитическим образом жизни, относят к дегенерации. А дегенерация, как и ароморфоз и идиоадаптация, тоже относится к биологическому процессу.
№ 7 Оба суждения неверны. А – генетический под триптолетен, т.е. в молекуле НК одна аминокислота кодируется сочетанием 3-х последовательно расположенных нуклеотидов.
Б – генетический код однозначен, т.е. один триплет (кодон) всегда кодирует только одну А.К.
№ 8 Верно только Б.
№ 10 Верны оба суждения
№ 14 Верны оба суждения
№ 16 Неверны оба суждения
Выберите книгу со скидкой:

Подарочный набор "Кисти и краски(4 по цене 2)"
350 руб. 297.00 руб.

Рисование головы и рук
350 руб. 1087.00 руб.

3D-рисование. Гиперреализм Рисунки, которые оживают
350 руб. 553.00 руб.

Изобразительное искусство.3 класс. Рабочий альбом
350 руб. 148.00 руб.

История цвета. Как краски изменили наш мир (новое оформление)
350 руб. 1025.00 руб.

350 руб. 1087.00 руб.

Радиевые девушки. Скандальное дело работниц фабрик, получивших дозу радиации от новомодной светящейся краски
350 руб. 427.00 руб.

Совушки. Раскраски, поднимающие настроение (ПР)
350 руб. 96.00 руб.

350 руб. 1087.00 руб.

Совушки. Раскраски, поднимающие настроение
350 руб. 283.00 руб.

Котики. Раскраски, поднимающие настроение
350 руб. 283.00 руб.

В цветочном вальсе. Открытки-раскраски
350 руб. 225.00 руб.
БОЛЕЕ 58 000 КНИГ И ШИРОКИЙ ВЫБОР КАНЦТОВАРОВ! ИНФОЛАВКА



- Карасаева Людмила ДзарахметовнаНаписать 0 20.11.2015
Номер материала: ДВ-174694
Добавляйте авторские материалы и получите призы от Инфоурок
Еженедельный призовой фонд 100 000 Р

Спикер: Анна Быкова (#лениваямама)
-
20.11.2015 789
-
20.11.2015 1387
-
20.11.2015 1872
-
20.11.2015 473
-
20.11.2015 2214
-
20.11.2015 1093
-
20.11.2015 618
Не нашли то что искали?
Вам будут интересны эти курсы:
| АТФ-синтазы | |
|---|---|
 |
| Поиск | |
|---|---|
| PMC | статьи |
| PubMed | статьи |
| NCBI | белки |
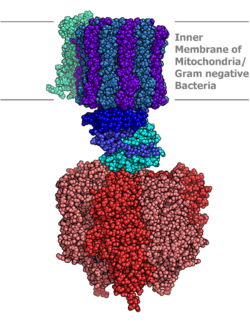
Образование АТФ из АДФ и P я энергетически невыгодно , и , как правило , протекают в обратном направлении. Для того , чтобы управлять этой реакцией вперед, АТФ - синтазы пар синтезом АТФ во время клеточного дыхания к электрохимическому градиенту , созданный разность протона (Н + ) концентрациям через митохондриальную мембрану в эукариоте или плазматической мембране бактерий. В ходе фотосинтеза в растениях, АТФ синтезируется АТФ - синтазы с использованием протонного градиента , созданный в тилакоидах просвета через тилакоидную мембрану и в хлоропласте стромы.
АТФ - синтаза состоит из двух основных субъединиц, F O и F 1 , который имеет вращательный механизм , позволяющий двигателя для производства АТФ. Из - за своей вращающейся субъединицы АТФ - синтазы является молекулярная машина .
содержание
Номенклатура
Структура и функции


Расположенный в пределах тилакоидной мембраны и внутренней митохондриальной мембраны , АТФ - синтаза состоит из двух областей F O и F 1 . Р О вызывает вращение F 1 и выполнен из с-кольца и субъединиц а, б, д, F6. F 1 состоит из субъединиц. Р 1 имеет водорастворимую часть , которая может гидролизовать АТФ. F O с другой стороны , имеет в основном гидрофобные области. F O F 1 создает путь для протонов движения через мембрану. α , β , γ , δ <\ Displaystyle \ альфа, \ бета, \ гамма, \ дельта>

F 1 часть АТФ - синтазы является гидрофильным и отвечает за гидролиз АТФ. Эта часть расположена через межмембранное пространство между мембраной среди пространства митохондрий. Субъединицы и сделать гексамеры с 6 сайтов связывания. Три из них являются каталитический неактивными , и они связывают АДФ. α <\ Displaystyle \ альфа>
β <\ Displaystyle \ бета>
Другие три субъединицы катализирует синтез АТФ. Другие F 1 субъединицы являются частью вращательного механизма двигателя. субъединица позволяет пройти через конформационные изменения, т.е. закрыт, наполовину открытые и открытые состояния позволяют АТФ быть связанными и выпустила сразу синтезируется. F 1 частица является большой и может рассматриваться в просвечивающем электронном микроскопе с помощью отрицательного окрашивания. Эти частицы диаметром 9 нм , что перец внутренней митохондриальной мембраны. γ , δ , ε <\ Displaystyle \ гамма, \ дельта, \ эпсилон>
γ <\ Displaystyle \ гамма>
β <\ Displaystyle \ бета>
| субблок | человек Gene |
|---|---|
| альфа | ATP5A1 , ATPAF2 |
| бета | ATP5B , ATPAF1 , C16orf7 |
| гамма | ATP5C1 |
| дельта | ATP5D |
| эпсилон | ATP5E |

Р О представляет собой нерастворимый в воде белок с восемью субъединиц и трансмембранного кольца. Кольцо имеет форму тетрамерную с спиралью белком петли спирали , которая идет хотя конформационные изменения , когда протонированных и депротонированных, толкая соседние субъединицы вращаться, вызывая вращение F O , который затем также влияет на конформацию F 1 , в результате переключения состояний альфа и бета - субъединицы. Р О область АТФ - синтазы является протон пор, внедренный в митохондриальной мембране. Она состоит из трех основных субъединиц, A, B, и С, и (у людей) шесть дополнительных субъединиц, д , е , F , г , F6 и 8 (или A6L). Атомная модель для димерных дрожжей F O области была определена крио-ЭМ при общем разрешение 3,6 Å. Эта часть фермента находится в митохондриальной внутренней мембране и пары протонов транслокации в вращение причины синтез АТФ в F 1 области.
| субблок | человек Gene |
|---|---|
| a | MT-ATP6 , MT-ATP8 |
| б | ATP5F1 |
| с | ATP5G1 , ATP5G2 , ATP5G3 |
Связывание модели
В 1960 - х годах через 1970 - х годов, Пол Бойер , А Калифорнийского университета профессор, разработал связывания изменения или флип-флоп, теорию механизма, который предположил , что синтез АТФ зависит от конформационных изменений в АТФ - синтазы , генерируемого при вращении гамма - субъединицей. Исследовательская группа John E. Walker , затем в MRC Лаборатории молекулярной биологии в Кембридже , кристаллизуется F 1 каталитический домен-АТФ - синтазы. Структура, в то время самой большой асимметричная структура белка известна, показали , что модель роторно-катализ Бойер был, по существу, правильно. Для выяснения этого Бойер и Уокер разделяют половину 1997 Нобелевской премии по химии .
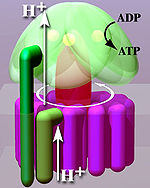
Кристаллическая структура F 1 показала переменный альфа- и бета - субъединицы (3 каждых), расположенную как дольки апельсина вокруг вращающейся асимметричной гаммы - субъединицы. В соответствии с текущей моделью синтеза АТФ (известной как переменная каталитической модель), трансмембранный потенциал , создаваемый (Н +) катионами протонов , поставляемых цепью переноса электронов, приводит в движении (H +) катионах протона из межмембранного пространства через мембрану с помощью F O область АТФ - синтазы. Часть F O (кольцо с-субъединиц ) вращается , как протоны проходят через мембрану. С-кольцо плотно прикреплено к асимметричной центральной ножке (состоящей в основном из гаммы - субъединицы), заставляя его вращаться внутри альфа - 3 беты 3 из F - вызывая 3 каталитических нуклеотидные сайты связывания , чтобы пройти через ряд конформационных изменений , которые привести к синтезу АТФ. Основные F 1 субъединицы предохранены от вращения в полном согласии с центральной ножкой ротором периферийного стеблом , который присоединяется к альфа - 3 беты 3 к невращающейся части F O . Структура неповрежденной АТФ - синтазы в настоящее время известно , при низком разрешении с электронной крио-микроскопии (крио-ЭМ) исследований комплекса. Модель крио-ЭМ АТФ - синтазы показывает , что периферийная Стебель представляет собой гибкую структуру , которая оборачивается вокруг комплекса , как он присоединяется F 1 до F O . При правильных условиях реакция фермента также может быть проведена в обратном направлении, при гидролизе АТФ движения протонов накачки через мембрану.
Физиологическая роль
Как и другие ферменты, активность F 1 F O АТФ - синтазы является обратимой. Широкоформатная достаточное количество АТФ заставляют его создавать трансмембранный протонный градиент , это используется брожение бактерий, не имеющих электронной транспортной цепи, а гидролизовать АТФ , чтобы сделать протонный градиент, который они используют для привода жгутиков и транспорт питательных веществ в клетку.
В дышащих бактерий в физиологических условиях, АТФ - синтазы, в общем, работает в противоположном направлении, создавая АТФ при использовании протонных движущей силы , созданной цепях переноса электронов в качестве источника энергии. Весь процесс создания энергии таким образом, называется окислительное фосфорилирование . Тот же самый процесс происходит в митохондриях , где АТФ - синтазы находятся во внутренней митохондриальной мембране , и Р 1 -часть проектах в митохондрии . Потребление АТФ АТФ-синтазы насосы катионов протонов в матрицу.
эволюция
Эволюция АТФ - синтазы , как полагают, был модульным , при которой два функционально независимых подразделения стали ассоциироваться и приобрел новые функциональные возможности . Эта ассоциация по- видимому, произошла в начале эволюционной истории, потому что по существу та же структура и активность АТФ - синтазы ферментов присутствует во всех царствах жизни. Синтазы показывает Р-АТФ с высоким функциональным и механистической сходство с V-АТФазы . Тем не менее, в то время как F-синтазы АТФ генерирует АТФ с использованием градиента протонов, то V-АТФазы генерирует протонный градиент за счет АТФ, генерации значений рН по цене от 1.
F 1 регион также показывает значительное сходство с гексамерным геликазом ДНК , а Р О области показывает некоторое сходство с Н +
Приведено жгутиковые моторные комплексы. Α 3 β 3 гексамеры из F 1 региона показывает значительное структурное сходство с гексамерным геликазом ДНК; и образуют кольцо с 3-кратным вращательной симметрии с центральным отверстием. Оба имеют роли , зависящие от относительного вращения макромолекулы внутри поры; в геликазах ДНК используют спиральную форму ДНК , чтобы управлять их движением вдоль молекулы ДНК и обнаружение суперспирализации, тогда как α 3 β 3 гексамеры используют конформационные изменения через вращение гаммы субъединицы для привода ферментативной реакции.
Н +
мотор F O частицы показывает большое функциональное сходство с H +
двигатели , которые управляют жгутиками. Оба имеют кольцо множества маленьких альфа-спиральных белков , которые вращаются относительно соседних стационарных белков, используя H +
градиент потенциала в качестве источника энергии. Эта ссылка является неопределенной, однако, как общая структура жгутиковых моторов гораздо сложнее , чем F O частицы и кольца с около 30 вращающихся белков намного больше , чем 10, 11 или 14 спиральных белков в F O сложный.
Модульная теория эволюции для возникновения АТФ - синтазы показывает , что две субъединицы с независимой функцией, ДНК геликазы с активностью АТФазы и H +
двигатель, способен связываться, а вращение двигателя вело АТФазы геликазов в обратном порядке . Этот комплекс затем эволюционировал большую эффективность и в конечном итоге превратился в сегодняшние сложные синтазы АТФ. Альтернативно, ДНК хеликаза / Н +
двигатель комплекс , возможно, имели H +
Активность насоса с активностью АТФазы геликазов , управляющих H +
двигатель в обратном направлении. Это, возможно, развились, чтобы осуществить обратную реакцию и выступать в качестве АТФ-синтазы.
У разных видов
E.coli , АТФ - синтаза является самым простым известной формой АТФ - синтазы, с 8 различных типов субъединиц.
Дрожжи АТФ - синтазы является одним из наиболее изученных эукариотических синтазы АТФ; и пять F 1 , восемь F O субъединиц, и семь ассоциированных белков были идентифицированы. Большинство из этих белков имеют гомологов в других эукариотов.
В растениях, АТФ - синтазы также присутствует в хлоропластах (CF 1 F O -АТФ - синтазы). Фермент интегрирован в тилакоидную мембрану; КВС 1 -часть палочки в строму , где темные реакции фотосинтеза (также называется свет независимых реакций или цикл Кальвина ) и синтез АТФ происходит. Общая структура и каталитический механизм синтазы хлоропласта АТФ почти такие же , как и у бактериального фермента. Тем не менее, в хлоропластах, то протонная движущая сила генерируется не дыхательной цепью переноса электронов , но первичными белками фотосинтетических.
АТФ - синтазы изолирован от крупного рогатого скота ( парнокопытных ) сердца митохондрий, с точки зрения биохимии и структуре, наиболее характеризующийся АТФ - синтазы. Говядина сердце используется в качестве источника для фермента из - за высокой концентрации митохондрий в сердечной мышце.
Ниже приведен список людей генов, которые кодируют компоненты синтазы АТФ:

Протонный потенциал движет бактерией
Они удивились еще больше, когда измерили скорость движения бактерий, обработанных арсенатом. Такая обработка снижала количество АТФ в клетке до практически неизмеримого уровня. И тем не менее лишенные АТФ бактерии отлично двигались, если в среде был кислород и протекал процесс дыхания.
Остановить бактерии удалось, добавив протонофор.
В то время концепция протонного потенциала была далеко не общепринятой даже в кругу биоэнергетиков. Поэтому вряд ли стоит удивляться, что микробиолог Адлер сформулировал свой вывод в рамках старой схемы, предполагавшей существование каких-то особых химических соединений, образуемых дыханием и потребляемых АТФ-синтетазой.
Однако для меня тогда уже было ясно, что у этих двух систем есть только один общий продукт — протонный потенциал. Стало быть, мутант кишечной палочки, исследованной американскими микробиологами, образовывал за счет дыхания протонный потенциал, который, по-видимому, и служил источником энергии для движения бактериальной клетки. Именно такое толкование опытов Адлера я предложил, выступая летом 1975 года на очередном съезде европейских биохимиков.
В подтверждение своей правоты я привел данные опытов, поставленных А. Глаголевым на пурпурной фотосинтезирующей бактерии. Испытывая различные комбинации ферментных ядов и разобщителей-протонофоров, Глаголев показал, что скорость движения микроба пропорциональна величине протонного потенциала, а не количеству АТФ. Это был важный шаг вперед по двум причинам.
Тем не менее нужен был прямой эксперимент. И он был вскоре поставлен.
Мы рассуждали таким образом. Если свет у нашей бактерии (или дыхание у кишечной палочки) нужен для движения только постольку, поскольку за их счет генерируется протонный потенциал, то можно получить подвижность и в отсутствие света (или дыхания), создав этот потенциал искусственно. Как это сделать?
Прежде всего необходимо перекрыть все пути образования протонного потенциала белками-генераторами. Затем к таким неподвижным уже бактериям надо добавить, например, кислоту, но не столько, чтобы, избави бог, их убить, а небольшое количество, которое просто создало бы некоторую избыточную концентрацию ионов водорода во внешней среде по сравнению с цитоплазмой бактериальной клетки. Поскольку в обычных условиях протонные генераторы бактерий откачивают ионы Н+ из клетки во внешнюю среду, то добавка кислоты должна имитировать включение генераторов.
С нетерпением я ждал результата этого опыта. Исполнится ли удивительное предсказание гипотезы: очнутся ли от паралича бактерии, отравленные целым коктейлем ядов, если в среду просто добавить немного соляной кислоты?
Удача? А может быть, капля кислоты просто разбавила яды?
Опыт повторяется, но вместо кислоты берется капелька воды. Нет эффекта: бактерии по-прежнему неподвижны. Еще один контроль: вместо кислоты добавляется щелочь. Эффекта нет и в этом случае.
А вдруг кислота разрушила какой-то из ядов? Это крайне маловероятно: ведь изменение кислотности среды в общем-то невелико. И тем не менее.
В работе с такой сложной системой, как живое существо, пусть даже мельчайшее, одноклеточное, всегда можно найти несколько объяснений любому факту. Однако стоящая гипотеза тем и хороша, что она не только объясняет старые факты, но и предсказывает новые. Именно так было установлено, что подвижность возвращается при добавке кислоты к отравленным ядами бактериям.
Известно, что время, необходимое, чтобы уравнялись концентрации ионов Н + между бактерией и средой, измеряется несколькими минутами. Значит, вызванное кислотой движение должно прекратиться спустя минуты.
И действительно, через три минуты после добавки кислоты поле под микроскопом являло собой печальную картину, которую мы наблюдали в начале опыта: бактерии были неподвижны.
Вот вам и разрушение яда кислотой! Что же это он сначала разрушился, а потом, когда кислота проникла в клетку, опять образовался?
А может быть, вообще клетка становится неподвижной, когда цитоплазма подкислилась?
Все может быть. Но заметьте, каждый следующий факт, предсказанный нашей гипотезой, требует от оппонента какого-нибудь нового предположения. Наша точка зрения ведет к новым фактам, противоположная — к новым предположениям.
И все же проверим, как влияет сама по себе кислотность среды на движение бактерий. Исключим из среды яд, мешавший производству протонного потенциала за счет света, и посмотрим, не обездвижутся ли бактерии при подкислении среды. Оказывается, этого не происходит. В подкисленной среде бактерии весело плавают до тех пор, пока не выключишь свет.
Если все это так, можно включить механизм движения и другим способом: создать, например, внутри клетки избыток отрицательных зарядов. Тогда даже при равенстве концентраций ионов Н + внутри и снаружи клетки эти ионы будут поступать внутрь за счет электрических сил, перемещаясь от плюса к минусу.
Сказано - сделано! На стекле две капли. В одной неподвижные, отравленные ядовитой смесью бактерии, в другой еще один яд, антибиотик валиномицин. Этот агент резко повышает проницаемость мембран для ионов калия (К + ).
Раствор валиномицина, как и среда с бактериями, не содержит ионов К + . В то же время внутри бактерий много этих ионов. Если теперь слить две капли, то валиномицин атакует бактерии, повысит их калиевую проницаемость и разрешит ионам К + выйти из бактерии, где их избыток по сравнению с окружающим раствором.
Выходя, ионы К + зарядят внутренность клетки отрицательно, этот минус притянет К + , и, двигаясь внутрь, Н+ запустит протонный мотор. Бактерии поплывут. Таково предсказание гипотезы.
А что получилось в опыте на самом деле? Бактерии задвигались и вновь через положенное время, когда уравнялись концентрации К+ внутри и снаружи клетки, остановились.
Предвидя новое возражение оппонента (а вдруг валиномицин работал у нас не переносчиком калия, а кем-то еще), мы поставили контрольный эксперимент, где бактерии находились в среде с высоким содержанием калия. Теперь калия было много и внутри и снаружи клетки. В таких условиях валиномицин не включал механизма подвижности. Эффект валиномицина (а также и кислоты) можно было снять и другим способом: добавив в среду разобщитель-протонофор и тем самым сведя к нулю протонный потенциал.
А совсем недавно Т, и А. Глаголевыми и М. Гусевым и К. Никитиной было доказано, что нитчатые сине-зеленые водоросли также используют протонный потенциал для своего скользящего движения по поверхности твердого субстрата.
Это последнее наблюдение свидетельствует, что протонный мотор, однажды изобретенный природой, применяется не только у бактерий, имеющих жгутики. У сине-зеленых водорослей жгутиков нет. Их роль выполняют, по-видимому, фибриллы, лежащие между внешней и цитоплазматической мембранами этих организмов, Сине-зеленые водоросли относятся к царству бактерий (у них есть даже другое название — цианобактерии). А могут ли организмы, принадлежащие к высшим царствам живой природы, двигаться за счет протонного потенциала? Чтобы ответить на этот вопрос, мы занялись движением хлоропластов.
Читайте также:
