
Особливості будови кори головного мозку
Архітектоніка кори - це загальний план будови кори. Загальна поверхня кори півкуль дорослої людини 2000 - 2500 см , причому 70% її заховані в глибині борозен. Товщина кори 2 - 4,5 мм. Йервові клітини і волокна, які утворюють кору, розташовані в 7 шарів:
- 1-й шар- молекулярний - найбільш поверхневий. У цьому шарі мало нервових клітин, вони дрібні. Шар утворений сплетінням нервових волокон. Завдяки цьому шару відбуваються внутрішньо- і міжпівкулеві зв'язки між різними частинами кори.
- 2-й шар- зовнішній зернистий. Складається з дрібних клітин у вигляді зернин і пірамід. Шар бідний на мієлінові волокна. Нейрони цього шару називаються вставними або інтернейронами. Ці нейрони забезпечують переробку інформації і її проведення до структур молекулярного шару на нижчі кіркові шари.
- 3-й шар- пірамідний, утворений середніми і великими пірамідними клітинами, з великою кількістю дендритів.
- 4-й шар- внутрішній зернистий, складається з дрібних зернистих клітин різної форми. Гранулярні нейрони, які розташовані у цьому шарі, здійснюють переробку і передачу інформації із закінчень аферентних волокон, які йдуть до кори і розгалужуються в межах I шару на пірамідні нейрони III і V шарів.
- 5-й шар- гангліозний - складається з великих пірамідних клітин. У передній центральній закрутці він містить ще "клітини Беца",аксони яких дають початок низхідним пірамідним шляхам, що проходять через стовбур головного мозку у спинний мозок і зв'язують кору півкуль з периферією. Аксони III і V шарів забезпечують різні види внутрішньо-кіркових, міжкіркових і кірково-підкіркових зв'язків. В цих шарах є також інтернейрони різних розмірів і форми, які забезпечують вибіркові внутрішньо-кіркові взаємодії між нейронами різних типів. Це необхідно для:
- - передачі інформації між вхідними в кору аферентними волокнами і пірамідними нейронами;
- - обміну інформацією між нейронами, які розташовані в різних кіркових шарах;
- - обміну інформацією між нейронами, які розташовані в різних звивинах, частках і півкулях;
- - зберігання і відтворення інформації (пам'ять).
- 6-й шар- поліморфний - складається з клітин трикутної і веретеноподібної форми і належать до білої речовини мозку.
- 7-й шар- складається з веретеноподібних нейронів, має багато волокон. Між нервовими клітинами всіх шарів виникають як постійні, так і тимчасові зв'язки.
Під корою міститься біла речовина півкуль мозку, в складі якої розрізняють асоціативні, комісуральні та проекційні волокна.
Асоціативні волокна зв'язують між собою окремі ділянки однієї півкулі. Короткі асоціативні волокна зв'язують між собою окремі закрутки і близькі поля, а довгі - закрутки різних часток у межах однієї півкулі.
Асоціативні поля беруть участь в інтеграції сенсорної інформації та забезпеченні зв'язків між чутливими й руховими зонами кори.
Асоціативні шляхи утворюються інтернейронами і їх волокнами. До асоціативних відносять також зв'язки, які утворюються між ядрами однієї половини стовбура мозку, проміжного мозку і базальними ядрами відповідної півкулі. У спинному мозку асоціативні нейрони забезпечують міжсегментарні зв'язки.
Комісуральні волокна зв'язують симетричні частини обох півкуль, більша частина проходить через мозолисте тіло.
Проекційні волокна виходять за межі півкуль, по них здійснюється двобічний зв'язок кори з відділами центральної нервової системи, що лежать нижче.
Проекційні шляхи можуть бути низхідними та висхідними.
Висхідні (сенсорні, чутливі або аферентні) проекційні шляхи проводять нервові імпульси від екстеро-, пропріо- і інтерорецепторів (чутливих нервових закінчень у шкірі, органів опорно-рухової системи, внутрішній органів), а також від органів чуття до головного мозку.
Крім кори головного мозку, сенсорна інформація може поступати і в інші відділи нервової системи, а саме, в мозочок, середній мозок, ретикулярну формацію.
Низхідні (еферентні) проекційні шляхи проводять нервові імпульси від кори великих півкуль до базальних і стовбурових ядер головного мозку, а потім до рухових ядер спинного мозку і стовбуру мозку. Вони передають інформацію, пов'язану з програмованим рухом організму в конкретних ситуаціях, тому є руховими провідними шляхами.
У товщі білої речовини півкуль є порожнини - бічні шлуночки, які протоками сполучаються з третім мозковим шлуночком.
У людини відомі випадки народження дітей, у яких немає кори великого мозку. Це - аненцефали. Вони живуть лише кілька днів. Все, що набувається організмом протягом індивідуального життя зв'язане з функцією великого мозку. З функцією кори великого мозку зв'язана вища нервова діяльність. Взаємодія організму із зовнішнім середовищем, його поведінка в навколишньому світі зв'язані з півкулями великого мозку.
Кора великих півкуль головного мозку (cortex cerebri) — це поверхневий шар сірої речовини. ЇЇ утворюють тіла нервових клітин, проміжки між якими заповнені клітинами нейроглії та містять нервові волокна і кровоносні судини. Наявність численних борозен і закруток значно збільшує площу сірої речовини. Загальна площа кори становить 220 тис. мм. Товщина кори в різних ділянках неоднакова і коливається від 1,3 до 4,5 мм. В ній міститься 14 — 15 млрд. різноманітних за формою, розмірами та функціями нейронів.
У різних тварин кора головного мозку розвинута неоднаково. В процесі еволюції кора виникла пізніше інших відділів нервової системи. Найбільш складну будову кора має у вищих ссавців. Особливого розвитку великі півкулі і кора досягли у людини. В міру розвитку кори підвищується значення її як вищого відділу нервової системи, що регулює функції організму і зумовлює зв’язок організму із зовнішнім середовищем, а також відіграє виключно важливе значення у здійсненні вищої нервової діяльності.
У корі головного мозку людини за сучасною класифікацією розрізняють давню (paleocortex), стару (archeocortex) і нову кору (neocortex), а також кору перехідного характеру — проміжну, або середню (mesocortex).
Давня кора виникла у процесі філогенезу в зв’язку з розвитком нюхового аналізатора (мозку). З виходом тварин на сушу почала інтенсивно розвиватися плащоподібна частина півкуль, відокремлена від підкірки білою речовиною і утворена старою і новою корою. У земноводних, поряд із давньою корою, з’являється зачаток старої кори, а у плазунів починає виникати нова кора, яка досягає найбільшого розвитку у ссавців, особливо у приматів.
У людини давньою корою називають невелику ділянку кори лобної частки, що розташована поблизу нюхової цибулини (передня дірчаста речовина), яка входить до системи нюхового аналізатора (мозку). Давня кора має три- та двошарову будову.
До старої кори відносяться гіпокамп, гачок парагіпокампової закрутки, поясна та зубчаста закрутки, що розміщені на медіальній поверхні півкуль, і мають лише три шари нейронів.
Проміжна кора займає незначну частину поверхні між старою і новою корою. До неї відноситься кора острівцевої частки та задня частина парагіпокампової закрутки.
Структури старої і проміжної кори входять до складу лімбічної системи, яка формує позитивні і негативні емоції та регулює вегетативні функції. Давня, стара і проміжна кора складає лише 4,4% поверхні великого мозку людини, а 95,6%, тобто переважна більшість його поверхні, становить нова кора.
50. Мікроскопічна будова мозкової кори.
Вивченням мікроскопічної будови кори займається наука, що називається архітектонікою кори великого мозку. Основними складовими розділами її є: цитоархітектоніка — вивчає особливості клітинної будови у різних ділянках та шарах кори; мієлоархітектоніка — вивчає особливості розміщення нервових волокон; ангіоархітектоніка — вивчає особливості будови та розташування кровоносних судин; синаптологія — вивчає сполучення між нервовими клітинами тощо. За цими ознаками в корі великого мозку розрізняють такі структурні одиниці, як зони, ділянки, поля та ін. Зараз відомо більше 200 цитоархітектонічних полів, що відрізняються своєю будовою.
Систематичне вивчення будови кори великого мозку почалося в кінці XIX століття. Одним із перших дослідників мікроскопічної будови кори був видатний український вчений професор Київського університету Володимир Олексійович Бец. Його праці мали особливо велике значення для розвитку цитоархітектоніки кори.
Кора великих півкуль, незважаючи на незначну товщину, має дуже складну будову. Починаючи з поверхні кори в глибину клітини утворюють декілька шарів, розміщених у певній послідовності, як це було встановлено В.О.Бецом ще в 1874 році. Залежно від філогенетичного походження окремі ділянки кори містять різну кількість шарів, які відрізняються будовою, формою, розмірами та розташуванням нервових клітин і волокон. Лише нова кора має типову шестишарову будову. Давня, стара і проміжна кора має значно простішу будову і містить значно меншу кількість в їхньому складі клітинних шарів.
Отже, в найбільш поширених ділянках кори розрізняють такі основні шари: перший — молекулярний, другий — зовнішній зернистий, третій — пірамідний, четвертий — внутрішній зернистий, п’ятий — гангліозний, шостий — шар поліморфних клітин. У деяких ділянках кори розрізняють і сьомий шар, що складається із веретеноподібних нейронів. Він містить порівняно з іншими шарами мало клітин і багато волокон.
Гангліозний шар клітин має в своєму складі гігантські пірамідні клітини (клітини Беца) вперше відкриті і описані професором В.О. Бецом. Нейрити цих клітин утворюють головну частину пірамідних провідних шляхів.
В кожному клітинному шарі, окрім нервових клітин, є ще й нервові волокна. Будова і щільність їх розташування також неоднакова в різних ділянках кори.
Між нервовими клітинами шарів кори в процесі їхньої діяльності існують як постійні, так і тимчасові зв’язки.
Кора представляет собой филогенетически наиболее молодой и вместе с тем сложный отдел мозга, предназначенный для обработки сенсорной информации, формирования поведенческих реакций организма.
Кора больших полушарий делится на древнюю (обонятельная луковица, обонятельный тракт, обонятельный бугорок), старую (часть лимбической системы) и новую кору. Новая кора занимает 95-96% общей площади и 4-5% приходится на долю древней и старой коры. Толщина коры колеблется от 1,3 до 4,5 мм. Площадь коры увеличивается за счет борозд и извилин. У взрослого человека она составляет 2200 см²
Кора состоит из серого и белого вещества, а также нейроглии. Количество нейронов 16-18 млрд. Глиальные клетки выполняют трофическую функцию.
По функциональному признаку нейроны коры делятся на 3 вида: афферентные (сенсорные) – к ним подходят нервные волокна афферентных путей, ассоциативные (вставочные) – в пределах головного и спинного мозга, эфферентные (двигательные) – образует нисходящие (эфферентные) проводящие пути, идущие от коры к разным ядрам головного и спинного мозга. К сенсорным клеткам относятся звездчатые клетки, входящие в 3 и 4 слоя сенсорных областей коры. К эфферентным нейронам относятся нейроны 5 слоя моторной зоны, которые представлены гигантскими пирамидными клетками Беца. К ассоциативным клеткам относятся веретенообразные и пирамидные клетки 3 слоя.
В связи с тем, что тела и отростки описанных выше нейронов имеют упорядоченное расположение, кора построена по экранному принципу, т.е. сигнал фокусируется не точка в точку, а на множество нейронов, что обеспечивает полный анализ раздражителя, а также возможность передачи сигнала в другие зоны коры, которые заинтересованы в нем.
Кора состоит из 7 слоев.
Молекулярный слой – мелкие нейроны и волокна. Сюда приходят афферентные таламокортикальные волокна от неспецифических ядер таламуса, регулирующие уровень возбудимости корковых нейронов.
Наружный зернистый слой образован мелкими нейронами в форме зерен и мелкими пирамидными клетками.
Наружный пирамидный слой состоит из пирамидных клеток разной величины. Функционально II и III слои коры объединяют нейроны, отростки которых обеспечивают кортико-кортикальные ассоциативные связи.
Внутренний зернистый слой образован звездчатыми клетками. Здесь оканчиваются афферентные таламокортикальные волокна, идущие от проекционных ядер таламуса.
Внутренний пирамидный слой включает крупные пирамидальные клетки – клетки Беца, аксоны которых идут в головной и спинной мозг.
Полиморфный слой ( мультиформный) – многоформенные нейроны, имеющие треугольную и веретенообразную форму.
Веретенообразные нейроны связывают все слои коры, их волокна поднимаются до 1 слоя. Имеются только в некоторых областях коры.
Функциональной единицей коры является вертикальная колонка, состоящая из 7 клеток, они вместе реагируют на один и тот же раздражитель.
В коре выделяют сенсорные, ассоциативные и двигательные зоны, исходя из расположения нейронов:
Сенсорные зоны – это входные участки коры, которые через восходящие нервные пути получают сенсорную информацию от большинства рецепторов тела.
Двигательные зоны – выходные области коры. В них возникают двигательные импульсы, идущие к произвольным мышцам по нисходящим путям, которые находятся в белом веществе больших полушарий.
Цитоархитектоника – это расположение нейронов в коре.
Миелоархитектоника – это распределение волокон в коре головного мозга.
Взаємовідносини борозен і звивини з кістками і швами черепа у новонародженої дитини інші, ніж у дорослого. Основні борозни (центральна, латеральна) виражені добре, але гілки основних борозен і дрібні звивини позначені слабо. Надалі в процесі розвитку кори борозни стають глибшими, а звивини між ними рельєфніше. Співвідношення борозен, звивин і швів черепа, характерне для дорослої людини, встановлюється у дітей в 6-8 років.
Протягом перших місяців життя розвиток кори йде дуже швидкими темпами. Більшість нейронів набувають зрілу форму, інтенсивно відбуваються процеси мієлінізації нервових волокон, що дозволяють реагувати на зовнішні подразники більш диференційовано.
У процесі еволюції людини як біологічного виду, а також у процесі онтогенезу - індивідуального розвитку кожної людини - відбувається кортікалізаціі функцій, тобто включення кори головного мозку в регуляцію функцій нижележащих структур мозку. Це дозволяє організувати більш досконалу, що враховує індивідуальний досвід, збережений у пам'яті, регуляцію функцій організму. Надалі, у міру автоматизації тієї чи іншої реакції, виконання її знову передається підкірковим структурам з формуванням автоматичного реагування.
Різні коркові зони дозрівають нерівномірно. Найбільш рано дозріває соматосенсорная і рухова кора, дещо пізніше - зорова і слухова. Особливо інтенсивним протягом першого півріччя життя є розвиток зорової кори, що тягне за собою розвиток інших зон мозку та їх інтеграцію. Дозрівання сенсорних і моторних зон в основному закінчується до 3 років. Значно пізніше дозріває асоціативна кора: до 7 років формуються її основні зв'язки, а остаточна диференціювання, формування нейронних ансамблів і зв`язків з іншими відділами мозку відбуваються до підліткового віку. Найбільш пізно (ближче до 9 років) дозрівають лобні області кори. Поступовість дозрівання структур кори великих півкуль визначає вікові особливості вищих нервових функцій і поведінкових реакцій дітей різних вікових груп.
Загальна площа кори головного мозку людини близько 2200 см2, число нейронів кори перевищує 10 млрд. У складі кори маються пірамідні, зірчасті, веретеноподібні нейрони.
Пірамідні нейрони мають різну величину, аксон пірамідного нейрона, як правило, проходить через білу речовину в інші зони кори або в інші мозкові структури.
Зірчасті клітини мають короткі добре гілкуються дендрити і короткий аксон, що забезпечує зв'язки нейронів в межах самої кори головного мозку.
Веретеноподібні нейрони забезпечують вертикальні або горизонтальні взаємозв'язку нейронів різних шарів кори.
Кора головного мозку має переважно шестіслойних будову (рис. 11.10).

Рис. 11.10. Будова кори головного мозку
Шар I - верхній молекулярний, представлений в основному ветвлениями висхідних дендритів пірамідних нейронів, серед яких розташовані рідкісні горизонтальні клітини і клітини-зерна, сюди ж приходять волокна неспецифічних ядер таламуса, що регулюють через дендрити цього шару рівень збудливості кори головного мозку.
Шар II - зовнішній зернистий, складається з зірчастих клітин, що визначають тривалість циркулювання збудження в корі головного мозку, тобто мають відношення до пам'яті.
Шар III - зовнішній пірамідний, формується з пірамідних клітин малої величини і разом з II шаром забезпечує Корко-коркові зв'язку різних звивин мозку.
Шар IV - внутрішній зернистий, містить переважно зірчасті клітини. Тут закінчуються специфічні таламокортікальние шляху, тобто шляху, що починаються від рецепторів аналізаторів.
Шар V - внутрішній пірамідний (гангліозний), шар великих пірамід, які є вихідними нейронами, аксони їх йдуть в стовбур мозку і спинний мозок. У руховій зоні в цьому шарі знаходяться гігантські пірамідні клітини, відкриті Бецом (клітини Беца).
Шар VI - шар поліморфних клітин, більшість нейронів цього шару утворюють кортико-таламические шляху.
Розподіл нейронів по шарах в різних областях кори дозволило виділити в мозку людини 53 цитоархитектонических поля (поля Бродмана), які вдосконалюються в міру розвитку кори головного мозку. У людини і вищих ссавців розрізняють, поряд з первинними, вторинні і третинні коркові поля, що забезпечують асоціацію функцій даного аналізатора з функціями інших аналізаторів.
Особливістю коркових полів є екранний принцип їх функціонування, що полягає в тому, що рецептор проектує свій сигнал не так на один нейрон кори, а на поле нейронів, яке утворюється їх зв'язками. У результаті сигнал фокусується не крапка в точку, а на безлічі різноманітних нейронів, що забезпечує його повний аналіз і можливість передачі в інші зацікавлені структури. Так, одне волокно, що приходить в зорову область кори, може активувати зону розміром 0,1 мм. Це означає, що один аксон розподіляє свою дію на більш ніж 5000 нейронів.
Функції окремих зон нової кори визначаються особливостями її структурної організації, зв'язками з іншими структурами мозку, участю в сприйнятті, зберіганні та відтворенні інформації при організації та реалізації поведінки, регуляції функцій сенсорних систем, внутрішніх органів.
Структурний відмінність ділянок кори головного мозку пов'язане з різницею їхніх функцій. У корі головного мозку виділяють сенсорні, моторні і асоціативні області (рис. 11.11).

Рис. 11.11. Цітоархітектоніческі поля Бродмана
Коркові кінці аналізаторів мають свою топографію - локальне розташування в певних ділянках кори головного мозку. Вони називаються сенсорними областями кори головного мозку. Коркові кінці аналізаторів різних сенсорних систем перекриваються. Крім цього, у кожній сенсорній системі кори маються полісенсорній нейрони, які реагують не тільки на "свій" адекватний стимул, але й на сигнали інших сенсорних систем. Ці механізми лежать в основі формування полімодальних зв'язків, що забезпечують сочетанную реакцію на різні подразники.
Шкірна рецептірующая система, таламокортікальние шляху проектуються на задню центральну звивину. Тут мається суворе соматотопическую поділ. На верхні відділи цієї звивини проектуються рецептивні поля шкіри нижніх кінцівок, на середні - тулуба, на нижні відділи - руки, голови.
На задню центральну звивину в основному проектується больова і температурна чутливість. У корі тім'яної частки (поля 5 і 7, див. Рис. 11.11), де також закінчуються провідні шляхи чутливості, здійснюється більш складний аналіз: локалізація роздратування, дискримінація, стереогіоз. При пошкодженнях кори особливо сильно порушуються функції дистальних відділів кінцівок, особливо рук.
Зорова система знаходиться в потиличній частці мозку: поля 17, 18, 19. Центральний зоровий шлях закінчується в поле 17; він інформує про наявність та інтенсивності зорового сигналу. У полях 18 і 19 аналізуються колір, форма, розміри, якість предметів. Поразка поля 19 кори головного мозку призводить до того, що хворий бачить, але не впізнає предмет (зорова агнозія, при цьому втрачається також колірна пам'ять).
Слухова система проектується в поперечних скроневих звивинах (звивини Гешля), в глибині задніх відділів латеральної (Сільвієвій) борозни (поля 41, 42, 52). Саме тут закінчуються аксони задніх горбів Четверохолміе і латеральних колінчастих тіл.
Нюхова система проектується в області переднього кінця гіппокампальних звивини (поле 34). Кора цій області має не шести-, а тришарове будову. При подразненні відзначаються нюхові галюцинації, пошкодження її веде до аносмія (втрати нюху).
Смакова система проектується в гіппокампальних звивині по сусідству з нюхової областю кори (поле 43).
У передній центральній звивині розташовані зони, подразнення яких викликає рух, вони представлені по соматотопическую типу, але зовсім інакше: у верхніх відділах звивини - нижні кінцівки, в нижніх - верхні. Це моторні області кори головного мозку.
Спереду від передньої центральної звивини лежать премоторні поля 6 і 8. Вони організовують не ізольовані, а комплексні, координовані, стереотипні рухи. Ці поля також забезпечують регуляцію тонусу гладкої мускулатури, пластичний тонус м'язів через підкіркові структури.
У реалізації моторних функцій беруть участь також друга лобова звивина, потилична, верхнетеменная області.
Рухова область кори як ніяка інша має велику кількість зв'язків з іншими аналізаторами, чим, мабуть, і зумовлена наявність в пий значного числа полісенсорних нейронів.
Всі сенсорні проекційні зони і моторні області кори займають менше 20% поверхні кори головного мозку. Решта - це асоціативні області. Кожна асоціативна область кори пов'язана потужними зв'язками з кількома проекційними областями. У асоціативних областях відбувається інтеграція разномодальних інформації, що дозволяє здійснювати усвідомлення надходить інформації та складні поведінкові акти. Асоціативні області мозку у людини найбільш виражені в лобової, тім'яної і скроневій.
Кожна проекційна область кори оточена асоціативними областями. Нейрони цих областей здатні до сприйняття разномодальних інформації, володіють великими здібностями до навчання. Полісенсорній нейронів асоціативної області кори забезпечує їх участь в об'єднанні, що надходить, забезпеченні взаємодії сенсорних і моторних областей кори.
Так, в тім'яній асоціативної області кори формуються суб'єктивні уявлення про навколишній простір, про нашому тілі. Це стає можливим завдяки зіставленню соматосенсорной, пропріоцептивної і зорової інформації. Лобові асоціативні поля мають зв'язку з лимбичним відділом мозку і беруть участь в організації програм дії при реалізації складних поведінкових актів з урахуванням їх емоційного забарвлення.
Першою і найбільш характерною рисою асоціативних областей кори є здатність їх нейронів сприймати разномодальную інформацію, причому сюди надходить не первинна, а вже оброблена інформація з виділенням біологічної значущості сигналу. Це дозволяє формувати програму цілеспрямованої поведінкового акту.
Друга особливість асоціативної області кори полягає в здатності до пластичних перебудовам в залежності від значимості надходить.
Третя особливість асоціативної області кори виявляється у тривалому зберіганні слідів сенсорних впливів. Руйнування асоціативної області призводить до виражених порушень навчання, пам'яті.
Розподіл функцій по областях мозку не є абсолютним. Встановлено, що практично всі області мозку мають полісенсорній нейрони, які певною мірою можуть брати на себе функцію пошкоджених модальноспсціфічсскіх нейронів. Це дозволяє компенсувати пошкодження структур мозку в ті періоди дитинства, коли пошкоджена функція ще не закріплена жорстко в структурі нервової тканини.
Важливою особливістю кори головного мозку є її здатність тривало зберігати сліди збудження. Це властивість надає корі виняткове значення в механізмах асоціативної переробки та зберігання інформації, накопичення знань.
Роль кори головного мозку в системній організації діяльності організму
1. Структурно-функціональні особливості кори
2. Роль кортикальних колонок
3. Проекційні та асоціативні поля
4. Методи вивчення кори великих півкуль
Кора великих півкуль з’являється вперше у риб у вигляді нюхових клітин, але видалення їх суттєво не впливає на поведінку риб. В амфібій вже є нюхова кора. Півкулі з’являються лише у птахів, у них при видаленні півкуль помітно змінюється поведінка: самі не можуть злетіти, самі не можуть їсти, в’ють гнізда, але не реагують на крик пташенят, у них зникає материнський інстинкт. Повного розвитку кора досягає у ссавців. У них видалення кори викликає втрату материнського інстинкту (собаки можуть поїдати своїх щенят), тварини не обходять перепони, натикаються на предмети, не йдуть на клич господаря, не орієнтуються в просторі, у них зникає захисний інстинкт. Найбільшу площу займає кора у приматів – 2200 см 2 . В них найкраще розвинені борозни та звивини. Функції кори повністю забезпечують пристосування до життя та вищу психічну діяльність. Видалення великих півкуль призводить до втрати здатності до самостійного життя.
У людини кора великих півкуль забезпечує такі функції:
- Взаємодія організму з навколишнім середовищем.
- Регуляція діяльності внутрішніх органів.
- Регуляція обміну речовин та енергії.
- Вища нервова діяльність –мова, пам’ять, мислення, свідомість.
Кора головного мозку є вищим відділом ЦНС. Це сіра речовина товщиною 3-5 мм, вкриває півкулі головного мозку. Вона займає площу 22 м 2 , утворюючи багаточисельні складки. В складі кори до 10 9 -10 10 нейронів, які утворюють 6 шарів:
1. Молекулярний шар – має мало клітин, їх волокна утворюють поверхневе густе тангенціальне сплетіння з дендритами другого шару.
2. Зовнішній зернистий шар – пірамідні клітини середньої величини, волокна яких розташовані радіально.
3. Внутрішній зернистий шар – зірчасті клітини, волокна яких розташовані горизонтально.
4. Внутрішній пірамідний (гангліозний) шар – це гігантські пірамідні клітини Беца, які мають довгі дендрити, що тягнуться до молекулярного шару.
5. Поліморфний шар – це шар веретеноподібних клітин.
Зв’язок кори великих півкуль з підкорковими структурами здійснюється за допомогою аферентних і еферентних волокон. Аферентні волокна називаються кортикопетальними , вони несуть інформацію в кору. Основними з них є таламокортикальн волокна . Це прямі аферентні шляхи, які розгалужуються у внутрішньому зернистому шарі і не дають колатералей. Невелика частина волокон йде в молекулярний шар, утворюючи колатеральні еферентні волокна, які називаються кортикофугальними , вони несуть інформацію від кори до підкоркових структур. Ці волокна діляться на 3 групи:
1. Проекційні – прямі еферентні волокна, що утворюють провідні шляхи (кортикоспінальні, кортикоталамічні)
2. Асоціативні - волокна, що утворюють безліч колатералей та йдуть в різні підкоркові зони однойменної півкулі.
3. Комісуральні - волокна, що йдуть від кори в складі мозолистого тіла і з’єднують зони кори однієї півкулі з підкорковими зонами другої.
1, 2 шари кори великих півкуль забезпечують аналіз та синтез отриманої інформації, мають багато асоціативних волокон.
3, 4 шари кори великих півкуль одержують інформацію від усіх органів та частин тіла за рахунок кортикопетальних волокон.
5, 6 шари кори великих півкуль – це рухові нейрони, звідси починаються рухові шляхи, що включають кортикофугальні волокна.
В шарах клітини розміщуються перпендикулярно до поверхні кори, утворюючи ланцюги. Елементарні нервові ланцюги відповідають за переробку певної інформації. Такий функціональний принцип названо – кортикальні колонки. Це елементарна функціональна одиниця, в якій здійснюється локальна переробка інформації від рецепторів однієї модальності. Кожна колонка має діаметр 500-1000 мкм, в складі яких розміщується 5-6 нейронів. Пірамідні клітини орієнтовані вертикально, їх аксони утворюють зворотні колатералі, які забезпечують як процеси полегшення в межах мікромодуля, так і гальмування між мікромодулями. Аксони зірчастих клітин ідуть через інтернейрони горизонтально, тому вони, головним чином, забезпечують гальмівні процеси. Веретеноподібні клітини мають довгі аксони, які орієнтовані як горизонтально, так і вертикально. Вони формують кортико-таламічні шляхи.
Мікромодулі об’єднуються в макромодулі завдяки горизонтальним розгалуженням терміналей. В колонці можуть бути прості та складні нейрони. Поряд з цим, в корі існує система, яка зчитує елементарні процеси в колонках та об’єднує всю інформацію. Формування таких систем зумовлено внутрішньо-кортикальними зв’язками між окремими макромодулями. Збудження одного мікромодуля викликає гальмування сусідніх. Активація мікромодулів відбувається за рахунок горизонтальних волокон таламокортикальних шляхів.
За функціями поля кори великих півкуль неоднозначні. Вивчення ролі окремих зон кори методом подразнення стало основою для вчення про локалізацію функцій в корі великих півкуль. Пізніше І.П. Павлов методом видалення в поєднанні з методом умовних рефлексів, підтвердив локалізацію функцій в корі. Разом з тим, Лешлі та ін. вважали, що різні зони кори великих півкуль є еквіпотенціальними, тобто однаковими за функціями – так виникла теорія еквіпотенціальності. Така думка була помилковою, бо тільки у нижчих тварин кора не має спеціалізованих зон. Так Лешлі, зруйнувавши кору щурів, знайшов, що порушення навиків руху щурів по лабіринту не залежить від місця руйнування. Досліди на тваринах з використанням мікроелектродного методу показали, що зони кори неоднозначні за функціями. Проекційні поля, створені мономодальними нейронами, одержують інформацію від рецепторів через переключаючі ядра таламуса. Вони оцінюють вид подразнення. На їх долю припадає 14% кори. Це специфічні проекційні поля. Асоціативні поля (86%) знаходяться навколо проекційних полів. Це полімодальні нейрони, які одержують інформацію від асоціативних ядер таламуса. Вони оцінюють джерело подразнення, його властивості та відстань до нього. Серед асоціативних полів є вищі поля, які забезпечують психічну діяльність людини, а також є поля, що формують пам’ять, індивідуальний досвід, прогнозування, передбачення. Це орбіто-фронтальні поля та тім’яна кора. Ці поля співпадають з функціональними зонами, де знаходяться нервові центри.
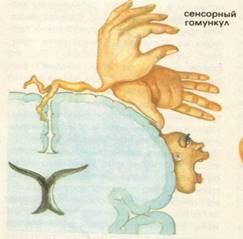
Представництво всіх видів чутливості в задній центральній звивині.
Задня центральна звивина – поля по Бродману – 1, 2, 3 – корковий центр всіх видів шкірної та пропріоцептивної чутливості. Вона має такі особливості:
1. Проекція протилежної сторони тіла розміщена вниз головою.
2. Має місце соматотопічна організація для кожної частини тіла.
3. Поля шкірної чутливості неоднозначні за площею, так, проекційні поля для обличчя, губ, кистей рук займають найбільшу площу.
При подразненні цієї зони виникає відчуття оніміння, тиску, повзання мурашок. При руйнуванні її зникає чутливість на протилежній стороні – анестезія.
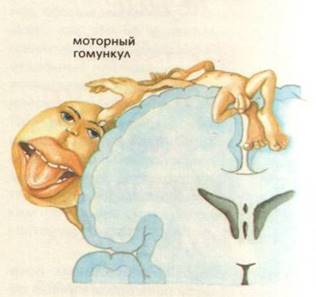
Проекція рухових центрів в передній центральній звивині.
Передня центральна звивина – поля по Бродману – 4, 6 – моторна зона. Вона забезпечує згинання, розгинання, пронацію, супінацію, ротацію. Вона також має соматотопічну організацію, але тут найбільшу площу займають мімічні м’язи обличчя, язик, кисті рук. При подразнення цієї зони виникає скорочення м’язів протилежної сторони тіла. При пошкодженні – розвиваються парези та паралічі. Парез – це обмеження довільних рухів. Параліч – це відсутність рухів.
Сенсорні системи мають подвійну організацію:
а) SS1 – це сомато-сенсорна зона – І, знаходиться в задній центральній звивині, виконує дискримінативний аналіз, тобто оцінює форму і характер поверхні предмету, приймає участь в складних рухах;
б) SS2 – це сомато-сенсорна зона – ІІ, має відношення до формування регуляції довільних рухів, орієнтації голови у напрямку звуків. Обидві зони мають сенсорні та моторні входи.
в) SМ1 – це сомато-моторна зона – І, знаходиться в передній центральній звивині;
г) SМ2 – це сомато-моторна зона – ІІ, знаходиться там же на медіальній поверхні кори. Обидві зони забезпечують відчуття просторової протяжності, стереогноз, відчуття ваги.
Потилична кора – це корковий центр зору. Він оцінює вигляд, форму, розміри та колір предмету. При пошкодження виникає геміанопсія (випадає частина поля зору), або виникає зорова агнозія (людина не впізнає знайомі предмети).
Верхня скронева звивина – корковий центр слуху (звивина Гешля – поля 41, 42). Оцінює звукові подразнення. При пошкодженні людина не може повторити сказане слово, хоч і чує, людина не розуміє звернену до неї мову, це слухова агнозія.
Задня частина нижньої лобної звивини (центр Брока) – це моторний центр мови. У правців він розміщується зліва. При пошкодження людина втрачає здатність до виразного мовлення, узгодження слів у реченні – моторна афазія.
Задня частина верхньої лобної звивини (центр Верніке) – це сенсорний центр мови, відповідає за сприйняття мови. При пошкодженні людина не розуміє звернену до неї мову – сенсорна афазія.
Задній відділ середньої лобної закрутки – руховий центр письма. При пошкодженні людина втрачає здатність писати – аграфія.
Кутова закрутка нижньої тім’яної долі (поле 30) – центр читання. При пошкодженні людина втрачає здатність читати – алексія.
Морський коник та його гачок – центр нюху. При пошкодженні людина втрачає здатність розрізняти запахи.
Верхні тім’яні долі – центр стереогнозу – оцінка положення тіла в просторі з закритими очима, глибока чутливість.
Надкрайова звивина (поле 40 лівої півкулі) – забезпечує здатність здійснювати привичні рухи чи автоматичні рухи, що сформувались в процесі життя. При пошкодженні розвивається моторна апраксія – людина знає, як виконати рух. але не може зробити його, не може намалювати предмет (порушується послідовність виконання складових руху).
Методи вивчення функцій кори великих півкуль:
1. Анатомо-клінічний метод – спостерігають за поведінкою людини при житті, вивчають структурні зміни після смерті.
2. Порівняльно-фізіологічний – оцінюють роль кори по поведінці у тварин різного рівня еволюційного розвитку.
3. Гістологічний – вивчають особливості будови кори на різних етапах розвитку організму.
4. Метод подразнення – вивчають функції кори шляхом подразнення різних її відділів.
5. Метод часткового та повного видалення кори – вивчають зміни поведінки тварин після видалення частин кори. (Роландо, 1823 рік).
6. Метод умовних рефлексів – І.П. Павлов вивчав поведінкові реакції у собак при сполученні різних подразників.
Генерацію електричних коливань в корі великого мозку живих організмів спостерігали і вперше описали Кеннон Р. (1875) і Данилевський В.Я. (1876). Реєстрація електричних коливань потенціалу від поверхні голови була проведена вперше в 1925 р. Правдич-Неминським В.В. у дослідах на тваринах і німецьким психіатром Н. Berger (1929) у людей.
У людей електрична активність кори великого мозку відводиться звичайно від шкіри на черепі. Метод реєстрації постійної електричної активності кори великого мозку отримав назву електроенцефалографія.
Для одержанная електроенцефалограми (ЕЕГ) можна застосувати або біполярний запис від двох активних електродів, розташованих на шкірі голови, або монополярний запис, при якому реєструючий (активний) електрод фіксують на шкірі голови, а індиферентний – на деякій відстані від нього (наприклад, на мочці вуха).
Запис ЕЕГ проводять за допомогою електроенцефалографу. При аналізі ЕЕГ враховують насамперед частоту, амплітуду, форму, тривалість та характер розподілу хвиль. Аналіз можна проводити візуально і за допомогою спеціальних електронних аналізаторів (аналогових і цифрових приладів). На сьогодні детально описано характеристики ЕЕГ для різних станів кори, точно виміряне і проаналізовано коливання потенціалів, які її складають. У несплячої людини в розслабленому стані при закритих очах за відсутності будь-яких зовнішніх подразників переважають хвилі частотою 8-13 Гц, амплітудою близько 50-100 мкВ, виражені вони більшою мірою в потиличних частках кори. Такі хвилі називають альфа-хвилями (або основним ритмом). Це так звана синхронізована ЕЕГ.
Коли людина розплющує очі, альфа-хвилі зникають (так звана блокада альфа-ритму або реакція десинхронізації) і замість них виникають бета-хвилі, які характеризуються більшою частотою (14-30 Гц) і меншою (до 30 мкВ) амплітудою. Вони відображають стан активності мозку, тобто збудження.
Під час перехідного стану від спокою до сну з’являється повільно-хвильовий ритм амплітудою 100-150 мкВ, частотою 4-7 Гц. Це тета-ритм. Під час глибокого сну на ЕЕГ реєструється дельта-ритм амплітудою 250-350 мкВ, частотою 0,5-4 Гц.
F - лобна кора, Р – тім’яна кора, О – потилична кора.
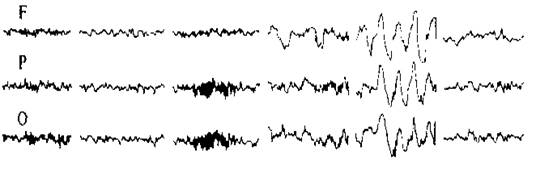
бета-ритм альфа-ритм тета-ритм дельта-ритм
ЕЕГ відображає головним чином постсинаптичні потенціали нейронів кори великого мозку. Про це свідчать результати одночасної реєстрації ЕЕГ за допомогою поза- і внутрішньоклітинного запису активності кортикальних нейронів. Припускають, що позитивні коливання потенціалів на поверхні кори великого мозку пов’язані з ЗПСП, який виникає в нейронах глибинних шарів кори або ГПСП у поверхневих її шарах.
Негативні коливання потенціалів ЕЕГ зумовлені зворотніми процесами в цих шарах. Ритмічна активність кори великого мозку, зокрема альфа-ритм, зумовлена головним чином впливами підкіркових структур, особливо таламуса, в якому виявлено особливі нейрони – “водії ритму” (пейсмекери), котрі через відповідні збудливі і гальмівні зв’язки здатні генерувати і підтримувати ритмічну активність кори великого мозку.
Запис ЕЕГ– діагностична процедура, яка використовується в неврологічній практиці. Так, при дифузних органічних пошкодженнях головного мозку, черепно-мозкових травмах спостерігаються сповільнені і нерегулярні хвилі. При пухлинах мозку часто виникають місцеві зміни ЕЕГ (у ділянці пухлин). У хворих на епілепсію на ЕЕГ спостерігаються пароксизмальні потенціали, судомні розряди, гострокінцеві хвилі та інші зміни. Запис ЕЕГ широко використовується в хірургічній практиці для контролю глибини наркозу: під час глибокої стадії наркотичного сну на ЕЕГ переважають дельта-хвилі.
При констатації смерті у сумнівних випадках, особливо при реанімації хворого, в клініці часто орієнтуються на зникнення коливань на ЕЕГ. (“плоска ЕЕГ”). У клінічній практиці також використовують метод реєстрації викликаних потенціалів для одержання об’єктивних даних про характер і динаміку деяких порушень сенсорних функцій.
При подразненні рецепторів шкіри, м’язів у первинних сомато-сенсорних ділянках переключаючих ядер таламуса кори великих півкуль реєструють коротко-латентні позитивно-негативні коливання електричного потенціалу, які носять назву первинні викликані потенціали. Вони постійні за формою, мають ЛП в межах 1-15 мсек, стійкі до наркозу та негативних чинників, здатні ритмічно відтворюватись. Зі значно більшим латентним періодом і більш складної форми реєструються потенціали, викликані подразненням неспецифічних висхідних шляхів, вони називаються вторинними потенціалами. Вторинні потенціали реєструються тільки в асоціативній корі, мають ЛП більше 15 мсек, зникають при поглибленні наркозу, нестабільні за формою, ритмічне відтворення їх одержати нелегко. Реєстрація викликаних потенціалів широко використовується для вивчення локалізації функцій в корі великих півкуль та встановлення зв’язків між різними структурами ЦНС.
Читайте также:
